
Plastid Phylogenomics Provide Evidence to Accept Two New Members of Ligusticopsis (Apiaceae, Angiosperms)

Abstract
:1. Introduction
2. Results
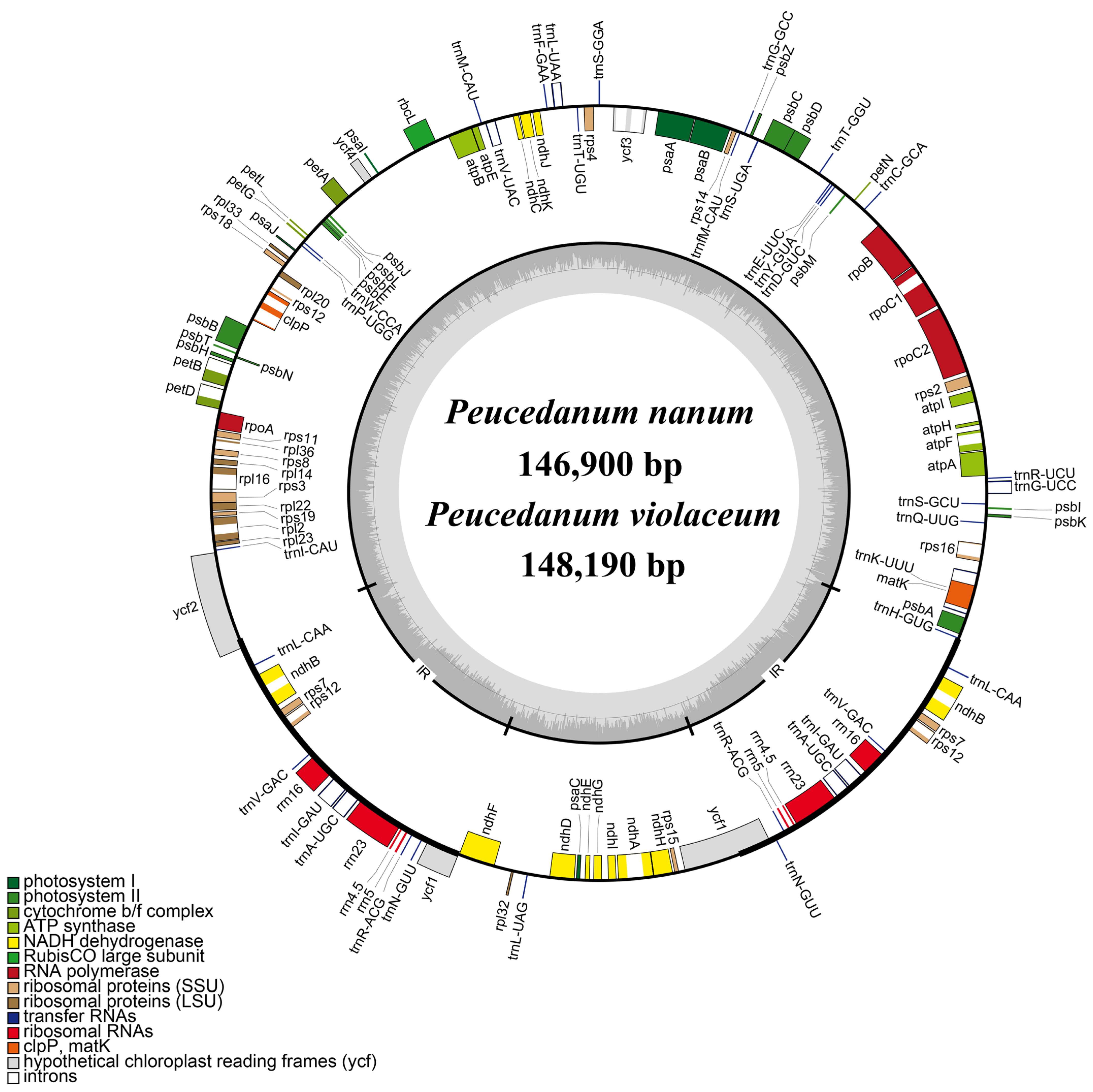
2.1. Plastome Features
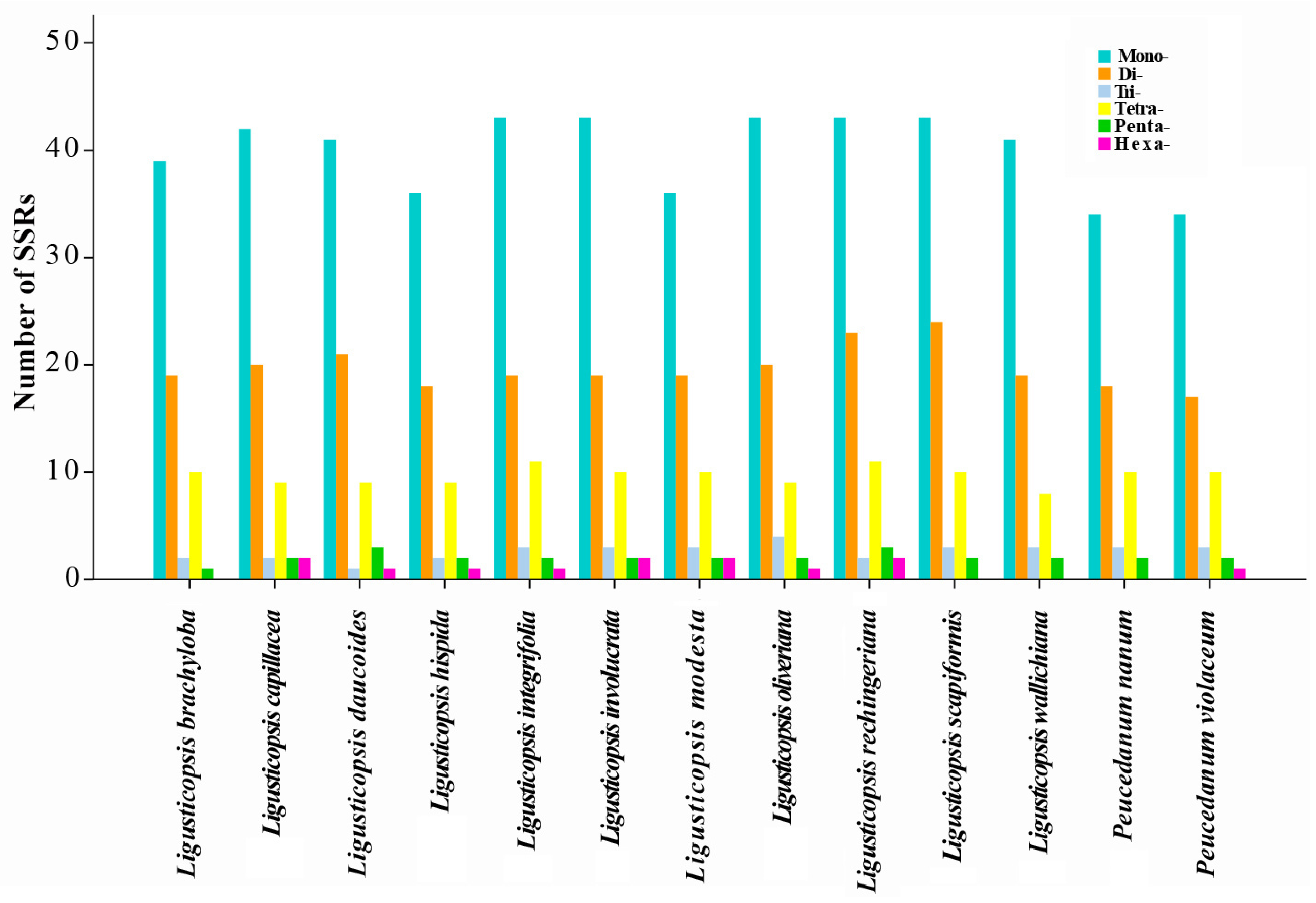
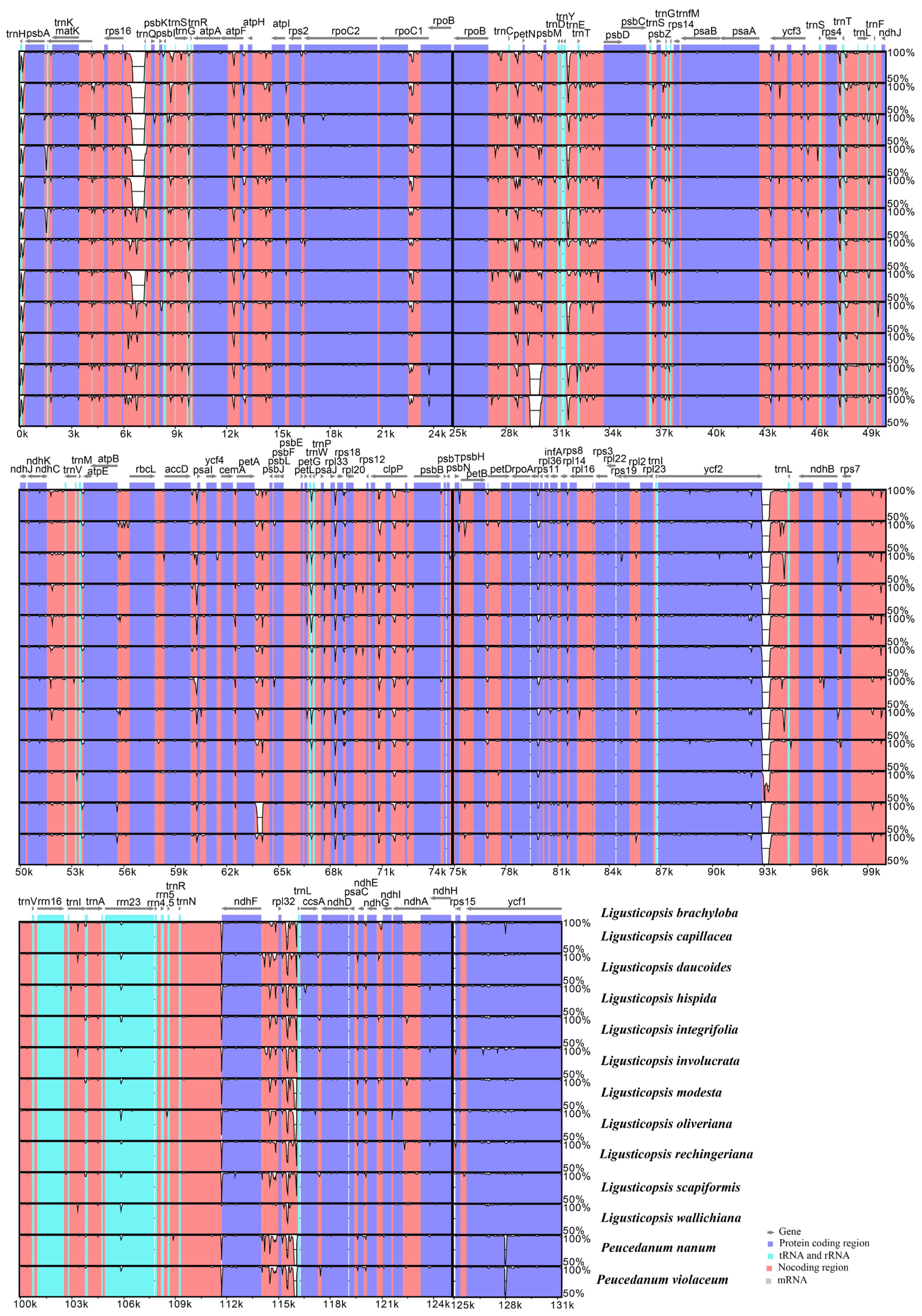
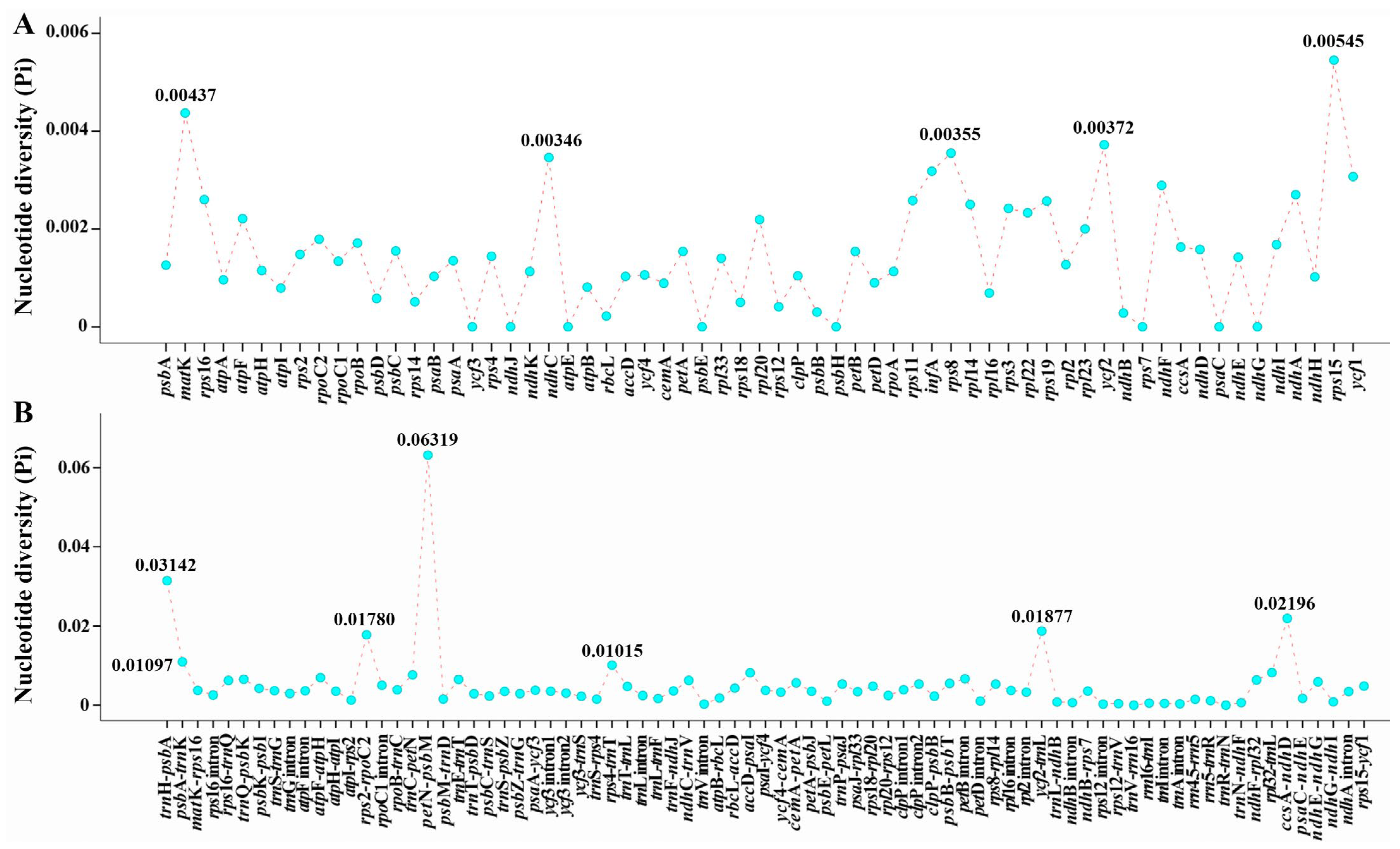
2.2. Plastome Comparison
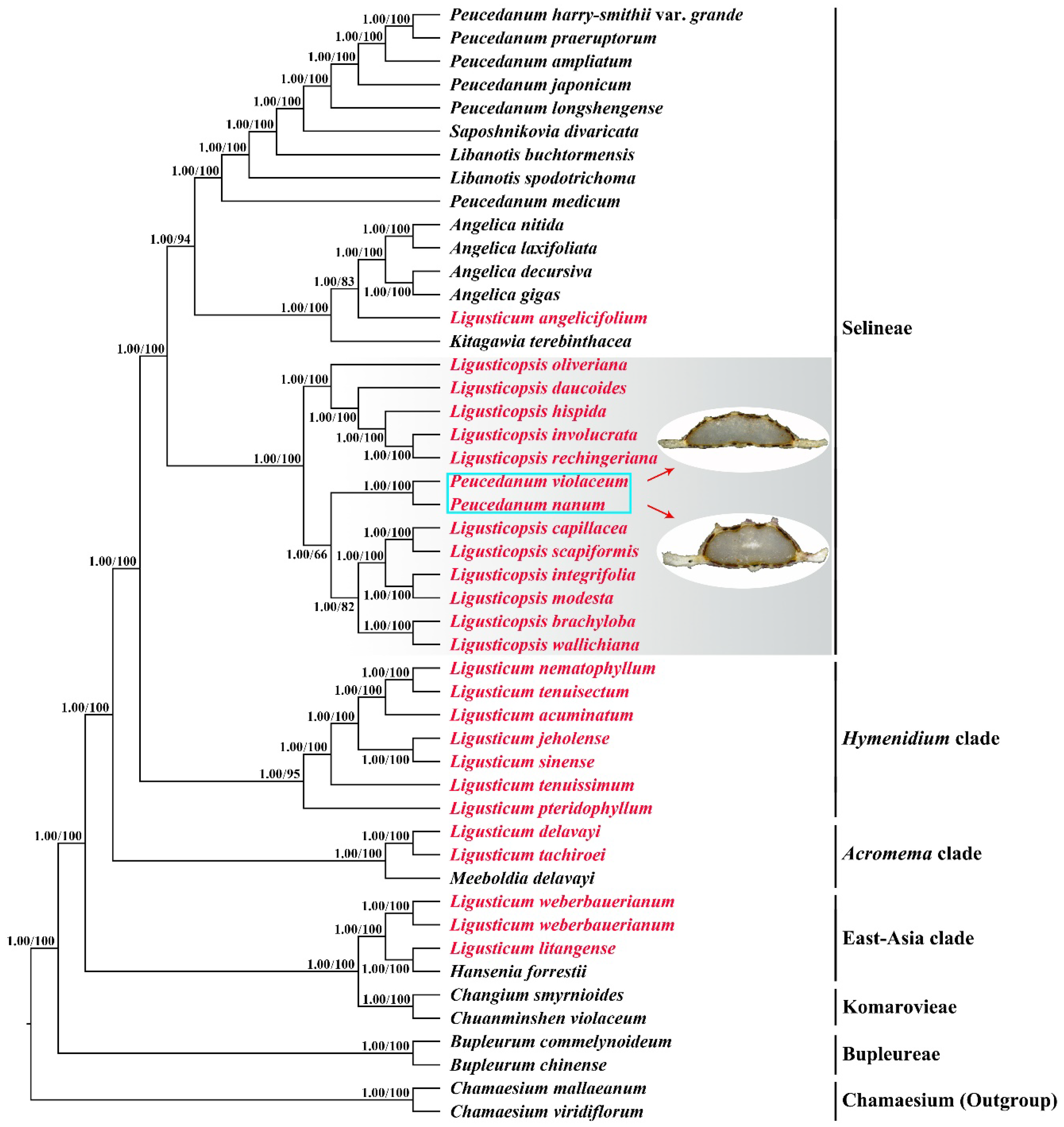
2.3. Phylogenetic Analyses
3. Discussion
3.1. Plastome Features
3.2. Phylogenetic Inference
3.3. Taxonomic Treatment
4. Materials and Methods
4.1. Plant Sample, DNA Extraction, Sequencing, and Assembly
4.2. Plastome Annotation and Feature Analyses
4.3. Comparative Plastome Analyses
4.4. Phylogenetic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Leute, G.H. Untersuchungen über den Verwandtschaftskreis der Gattung Ligusticum L. (Umbelliferae)—I Teil. Ann. Naturhist. Mus. Wien 1969, 73, 55–98. [Google Scholar]
- Pu, F.D. A revision of the genus Ligusticum L. (Umbelliferae) in China. Acta Phytotax. Sin. 1991, 29, 385–393. [Google Scholar]
- Leute, G.H. Untersuchungen über den Verwandtschaftskreis der Gattung Ligusticum L. (Umbelliferae)—II Teil. Ann. Naturhist. Mus. Wien 1970, 74, 457–519. [Google Scholar]
- Zhang, H.C. Ligusticum L. In Flora Reipublicae Popularis Sinicae; Shan, R.H., She, M.L., Eds.; Science Press: Beijing, China, 1985; Volume 55, pp. 234–257. [Google Scholar]
- Pu, F.D.; Watson, M.F. Ligusticum L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2005; Volume 14, pp. 140–150. [Google Scholar]
- Sun, N.; He, X.J.; Zhou, S.D. Morphological cladistic analysis of Ligusticum (Umbelliferae) in China. Nord. J. Bot. 2008, 26, 118–128. [Google Scholar] [CrossRef]
- Li, Z.X.; Guo, X.L.; Price, M.; Zhou, S.D.; He, X.J. Phylogenetic position of Ligusticopsis (Apiaceae, Apioideae): Evidence from molecular data and carpological characters. AoB Plants 2022, 14, plac008. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Xie, D.; Peng, C.; Gui, L.; Price, M.; Zhou, S.D.; He, X.J. Molecular evolution and phylogenetic relationships of Ligusticum (Apiaceae) inferred from the whole plastome sequences. BMC Eco. Evol. 2022, 22, 55. [Google Scholar] [CrossRef] [PubMed]
- Sheh, M.L. Peucedanum L. In Flora Reipublicae Popularis Sinica; Shan, R.H., Sheh, M.L., Eds.; Science Press: Beijing, China, 1992; Volume 55, pp. 123–175. [Google Scholar]
- Sheh, M.L.; Watson, M.F. Peucedanum L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2005; Volume 14, pp. 182–192. [Google Scholar]
- Shan, R.H.; Sheh, M.L.; Yuan, C.Q.; Wang, T.S. New taxa of the Umbelliferae from Xizang (Tibet). Acta Phytotax. Sin. 1980, 18, 374–379. [Google Scholar]
- Kadereit, J.W.; Bittrich, V. Flowering plants. Eudicots: Apiales, Gentianales (except Rubiaceae). In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 15, p. 168. [Google Scholar]
- Downie, S.R.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S. Molecular systematics of Old World Apioideae (Apiaceae): Relationships among some members of tribe Peucedaneae sensu lato, the placement of several island-endemic species, and resolution within the apioid superclade. Can. J. Bot. 2000, 78, 506–528. [Google Scholar]
- Spalik, K.; Reduron, J.P.; Downie, S.R. The phylogenetic position of Peucedanum sensu lato and allied genera and their placement in tribe Selineae (Apiaceae, subfamily Apioideae). Plant Syst. Evol. 2004, 243, 189–210. [Google Scholar] [CrossRef]
- Valiejo-Roman, C.M.; Terentieva, E.I.; Samigullin, T.H.; Pimenov, M.G.; Ghahremani-Nejad, F.; Mozaffarian, V. Molecular data (nrITS-sequencing) reveal relationships among Iranian endemic taxa of the Umbelliferae. Feddes Repert. 2006, 117, 367–388. [Google Scholar] [CrossRef]
- Feng, T.; Downie, S.R.; Yu, Y.; Zhang, X.M.; Chen, W.W.; He, X.J.; Liu, S. Molecular systematics of Angelica and allied genera (Apiaceae) from the Hengduan Mountains of China based on nrDNA ITS sequences: Phylogenetic affinities and biogeographic implications. J. Plant Res. 2009, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gong, X.; Downie, S.R.; Peng, H. Towards a more robust molecular phylogeny of Chinese Apiaceae subfamily Apioideae: Additional evidence from nrDNA ITS and cpDNA intron (rpl16 and rps16) sequences. Mol. Phylogenet. Evol. 2009, 53, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Gao, Y.Z.; Wei, J.; Liu, Z.W.; Downie, S.R. Molecular phylogenetics of Ligusticum (Apiaceae) based on nrDNA ITS sequences: Rampant polyphyly, placement of the Chinese endemic species, and a much-reduced circumscription of the genus. Int. J. Plant Sci. 2020, 181, 306–323. [Google Scholar] [CrossRef]
- Liu, C.K.; Lei, J.Q.; Jiang, Q.P.; Zhou, S.D.; He, X.J. The complete plastomes of seven Peucedanum plants: Comparative and phylogenetic analyses for the Peucedanum genus. BMC Plant Biol. 2022, 22, 101. [Google Scholar] [CrossRef]
- Shaw, J.; Shafer, H.L.; Leonard, O.R.; Kovach, M.J.; Schorr, M.; Morris, A.B. Chloroplast DNA sequence utility for the lowest phylogenetic and phylogeographic inferences in angiosperms, the tortoise and the hare IV. Am. J. Bot. 2014, 101, 1987–2004. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, X.; Yang, Z.; Yang, C.; Yang, J.; Ji, Y. Analysis of complete chloroplast genome sequences improves phylogenetic resolution in Paris (Melanthiaceae). Front. Plant Sci. 2016, 7, 1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonti-Filippini, J.; Nevill, P.G.; Dixon, K.; Small, I. What can we do with 1000 plastid genomes? Plant J. 2017, 90, 808–818. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Yang, L.; Chase, M.W.; Liu, C.; Yang, Z.; Yang, J.; Yang, J.B.; Yi, T.S. Plastome phylogenomics, biogeography, and clade diversification of Paris (Melanthiaceae). BMC Plant Biol. 2019, 19, 543. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Liu, Y.; Xu, C.; Gao, Y.; Yuan, Q.; Suo, Z.; Zhang, Z.; Sun, J. Chloroplast phylogenomic insights into the evolution of Distylium (Hamamelidaceae). BMC Genom. 2021, 22, 293. [Google Scholar] [CrossRef]
- Guo, X.; Liu, C.; Wang, H.; Zhang, G.; Yan, H.; Jin, L.; Su, W.; Ji, Y. The complete plastomes of two flowering epiparasites (Phacellaria glomerata and P. compressa): Gene content, organization, and plastome degradation. Genomics 2021, 113, 447–455. [Google Scholar] [CrossRef]
- Odago, W.O.; Waswa, E.N.; Nanjala, C.; Mutinda, E.S.; Wanga, V.O.; Mkala, E.M.; Oulo, M.A.; Wang, Y.; Zhang, C.F.; Hu, G.W.; et al. Analysis of the complete plastomes of 31 Species of Hoya group: Insights into their comparative genomics and phylogenetic relationships. Front. Plant Sci. 2021, 12, 814833. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.V.; Paule, J.; Jungcurt, T.; Cardoso, D.; Amorim, A.M.; Berberich, T.; Zizka, G. Resolving recalcitrant clades in the pantropical Ochnaceae: Insights from comparative phylogenomics of plastome and nuclear genomic data derived from targeted sequencing. Front. Plant Sci. 2021, 12, 638650. [Google Scholar] [CrossRef] [PubMed]
- Sielemann, K.; Pucker, B.; Schmidt, N.; Viehöver, P.; Weisshaar, B.; Heitkam, T.; Holtgräwe, D. Complete pan-plastome sequences enable high resolution phylogenetic classification of sugar beet and closely related crop wild relatives. BMC Genom. 2022, 23, 113. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Fu, J.; Fang, Y.; Xiang, J.; Dong, H. Complete chloroplast genomes of Rubus species (Rosaceae) and comparative analysis within the genus. BMC Genom. 2022, 23, 32. [Google Scholar] [CrossRef]
- Liu, C.; Yang, Z.; Yang, L.; Yang, J.; Ji, Y. The complete plastome of Panax stipuleanatus: Comparative and phylogenetic analyses of the genus Panax (Araliaceae). Plant Divers. 2018, 40, 265–276. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Z.; Liu, C.; He, Z.; Zhang, Z.; Yang, J.; Liu, H.; Yang, J.; Ji, Y. Chloroplast phylogenomic analysis provides insights into the evolution of the largest eukaryotic genome holder, Paris japonica (Melanthiaceae). BMC Plant Biol. 2019, 19, 293. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Yang, J.; Jin, L.; Wang, S.; Yang, Z.; Ji, Y. Plastome phylogenomics of the East Asian endemic genus Dobinea. Plant Divers. 2021, 43, 35–42. [Google Scholar] [CrossRef]
- Asaf, S.; Ahmad, W.; Al-Harrasi, A.; Khan, A.L. Uncovering the first complete plastome genomics, comparative analyses, and phylogenetic dispositions of endemic medicinal plant Ziziphus hajarensis (Rhamnaceae). BMC Genom. 2022, 23, 83. [Google Scholar] [CrossRef]
- Cvetković, T.; Hinsinger, D.D.; Thomas, D.C.; Wieringa, J.J.; Velautham, E.; Strijk, J.S. Phylogenomics and a revised tribal classification of subfamily Dipterocarpoideae (Dipterocarpaceae). Taxon 2022, 71, 85–102. [Google Scholar] [CrossRef]
- Xia, M.Z.; Li, Y.; Zhang, F.Q.; Yu, J.Y.; Khan, G.; Chi, X.F.; Xu, H.; Chen, S.L. Reassessment of the phylogeny and systematics of Chinese Parnassia (Celastraceae): A thorough investigation using whole plastomes and nuclear ribosomal DNA. Front. Plant Sci. 2022, 13, 855944. [Google Scholar] [CrossRef]
- Guo, X.L.; Zheng, H.Y.; Price, M.; Zhou, S.D.; He, X.J. Phylogeny and comparative analysis of Chinese Chamaesium species revealed by the complete plastid genome. Plants 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Gou, W.; Jia, S.B.; Price, M.; Guo, X.L.; Zhou, S.D.; He, X.J. Complete plastid genome sequencing of eight species from Hansenia, Haplosphaera and Sinodielsia (Apiaceae): Comparative analyses and phylogenetic implications. Plants 2020, 9, 1523. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, X.; Sun, J.; Wang, Y.; Ge, Y.; Dong, W.; Yuan, Q.; Huang, L. Phylogenomic and evolutionary dynamics of inverted repeats across Angelica plastomes. BMC Plant Biol. 2021, 21, 26. [Google Scholar] [CrossRef]
- Wen, J.; Xie, D.F.; Price, M.; Ren, T.; Deng, Y.Q.; Gui, L.J.; Guo, X.L.; He, X.J. Backbone phylogeny and evolution of Apioideae (Apiaceae): New insights from phylogenomic analyses of plastome data. Mol. Phylogenet. Evol. 2021, 161, 107183. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.P.; Liu, C.K.; Xie, D.F.; Zhou, S.D.; He, X.J. Plastomes provide insights into differences between morphology and molecular phylogeny: Ostericum and Angelica (Apiaceae) as an example. Diversity 2022, 14, 776. [Google Scholar] [CrossRef]
- Yang, L.; Abduraimov, O.; Tojibaev, K.; Shomurodov, K.; Zhang, Y.M.; Li, W.J. Analysis of complete chloroplast genome sequences and insight into the phylogenetic relationships of Ferula L. BMC Genom. 2022, 23, 643. [Google Scholar] [CrossRef]
- Ren, T.; Li, Z.X.; Xie, D.F.; Gui, L.J.; Peng, C.; Wen, J.; He, X.J. Plastomes of eight Ligusticum species: Characterization, genome evolution, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 519. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M. Refining the DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19451–19452. [Google Scholar] [CrossRef] [Green Version]
- CBOL Plant Working Group. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar] [CrossRef] [Green Version]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Li, D.Z.; van der Bank, M.; Twyford, A.D. Telling plant species apart with DNA: From barcodes to genomes. Philos. Trans. R. Soc. B 2016, 371, 20150338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaskin, J.F.; Wilson, L.M. Phylogenetic relationships among native and naturalized Hieracium (Asteraceae) in Canada and the United States based on plastid DNA sequences. Syst. Bot. 2007, 32, 478–485. [Google Scholar] [CrossRef]
- Theis, N.; Donoghue, M.J.; Li, J. Phylogenetics of the Caprifolieae and Lonicera (Dipsacales) based on nuclear and chloroplast DNA sequences. Syst. Bot. 2008, 33, 776–783. [Google Scholar] [CrossRef]
- Yue, J.P.; Sun, H.; Baum, D.A.; Li, J.H.; Al-shehbaz, I.A.; Pee, R. Molecular phylogeny of Solms-Laubachia (Brassicaceae) s.l., based on multiple nuclear and plastid DNA sequences, and its biogeographic implications. J. Syst. Evol. 2009, 47, 402–415. [Google Scholar] [CrossRef]
- Huang, J.L.; Sun, G.L.; Zhang, D.M. Molecular evolution and phylogeny of the angiosperm ycf2 gene. J. Syst. Evol. 2010, 48, 240–248. [Google Scholar] [CrossRef]
- Nakaji, M.; Tanaka, N.; Sugawara, T. A molecular phylogenetic study of Lonicera L. (Caprifoliaceae) in Japan based on chloroplast DNA sequences. APG 2015, 66, 137–151. [Google Scholar]
- Liu, P.L.; Wen, J.; Duan, L.; Arslan, E.; Ertuğrul, K.; Chang, Z.Y. Hedysarum L. (Fabaceae: Hedysareae) is not monophyletic–evidence from phylogenetic analyses based on five nuclear and five plastid sequences. PLoS ONE 2017, 12, e0170596. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.O.; Vieira, L.N.; Stefenon, V.M.; Faoro, H.; Pedrosa, F.O.; Guerra, M.P.; Nodari, R.O. Molecular relationships of Campomanesia xanthocarpa within Myrtaceae based on the complete plastome sequence and on the plastid ycf2 gene. Genet. Mol. Biol. 2020, 43, e20180377. [Google Scholar] [CrossRef]
- Wang, P.L.; Pu, F.D.; Ma, J.S. Pollen morphology of the genus Ligusticum from China and its systematic significance. Acta Phytotax. Sin. 1991, 29, 235–245. [Google Scholar]
- Pimenov, M.G.; Kljuykov, E.V.; Ostroumova, T.A. Himalayan species of Selinum L. s.l. (Umbelliferae). The genus Oreocome Edgew. Willdenowia 2001, 31, 101–124. [Google Scholar] [CrossRef]
- Pimenov, M.G. Updated checklist of Chinese Umbelliferae: Nomenclature, synonymy, typification, distribution. Turczaninowia 2017, 20, 106–239. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; de Pamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an integrated plastome sequence annotator and analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Mower, J.P. The PREP suite: Predictive RNA editors for plant mitochondrial genes, chloroplast genes, and user-defined alignments. Nucleic Acids Res. 2009, 37, W253–W259. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbech, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Total Length (bp) | LSC (bp) | SSC (bp) | IR (bp) | Total GC Content (%) | Total Genes (Unique) | Protein Coding Genes (Unique) | rRNA Genes (Unique) | tRNA Genes (Unique) |
|---|---|---|---|---|---|---|---|---|---|
| L. brachyloba | 148,633 | 92,265 | 17,588 | 19,390 | 37.40% | 113 | 79 | 4 | 30 |
| L. capillacea | 147,808 | 91,907 | 17,503 | 19,199 | 37.50% | 113 | 79 | 4 | 30 |
| L. daucoides | 148,078 | 91,666 | 17,582 | 19,415 | 37.40% | 113 | 79 | 4 | 30 |
| L. hispida | 147,797 | 91,846 | 17,627 | 19,162 | 37.40% | 113 | 79 | 4 | 30 |
| L. integrifolia | 148,196 | 92,305 | 17,575 | 19,158 | 37.50% | 113 | 79 | 4 | 30 |
| L. involucrata | 147,752 | 91,782 | 17,560 | 19,205 | 37.40% | 113 | 79 | 4 | 30 |
| L. modesta | 148,133 | 92,247 | 17,568 | 19,159 | 37.50% | 113 | 79 | 4 | 30 |
| L. oliveriana | 148,175 | 92,273 | 17,534 | 19,184 | 37.50% | 113 | 79 | 4 | 30 |
| L. rechingeriana | 148,525 | 91,813 | 17,654 | 19,529 | 37.30% | 113 | 79 | 4 | 30 |
| L. scapiformis | 148,107 | 92,214 | 17,581 | 19,156 | 37.50% | 113 | 79 | 4 | 30 |
| L. wallichiana | 148,594 | 92,281 | 17,567 | 19,373 | 37.40% | 113 | 79 | 4 | 30 |
| P. nanum | 146,900 | 91,480 | 17,308 | 19,056 | 37.50% | 113 | 79 | 4 | 30 |
| P. violaceum | 148,190 | 91,811 | 16,335 | 20,022 | 37.50% | 113 | 79 | 4 | 30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Deng, J.; Zhou, R.; Song, B.; Zhou, S.; He, X. Plastid Phylogenomics Provide Evidence to Accept Two New Members of Ligusticopsis (Apiaceae, Angiosperms). Int. J. Mol. Sci. 2023, 24, 382. https://doi.org/10.3390/ijms24010382
Liu C, Deng J, Zhou R, Song B, Zhou S, He X. Plastid Phylogenomics Provide Evidence to Accept Two New Members of Ligusticopsis (Apiaceae, Angiosperms). International Journal of Molecular Sciences. 2023; 24(1):382. https://doi.org/10.3390/ijms24010382
Chicago/Turabian StyleLiu, Changkun, Jiaojiao Deng, Renxiu Zhou, Boni Song, Songdong Zhou, and Xingjin He. 2023. "Plastid Phylogenomics Provide Evidence to Accept Two New Members of Ligusticopsis (Apiaceae, Angiosperms)" International Journal of Molecular Sciences 24, no. 1: 382. https://doi.org/10.3390/ijms24010382