
Plastid Phylogenomic Analyses Reveal a Cryptic Species of Ligusticopsis (Apiaceae, Angiosperms)

Abstract
:1. Introduction
2. Results
2.1. Morphological Analysis
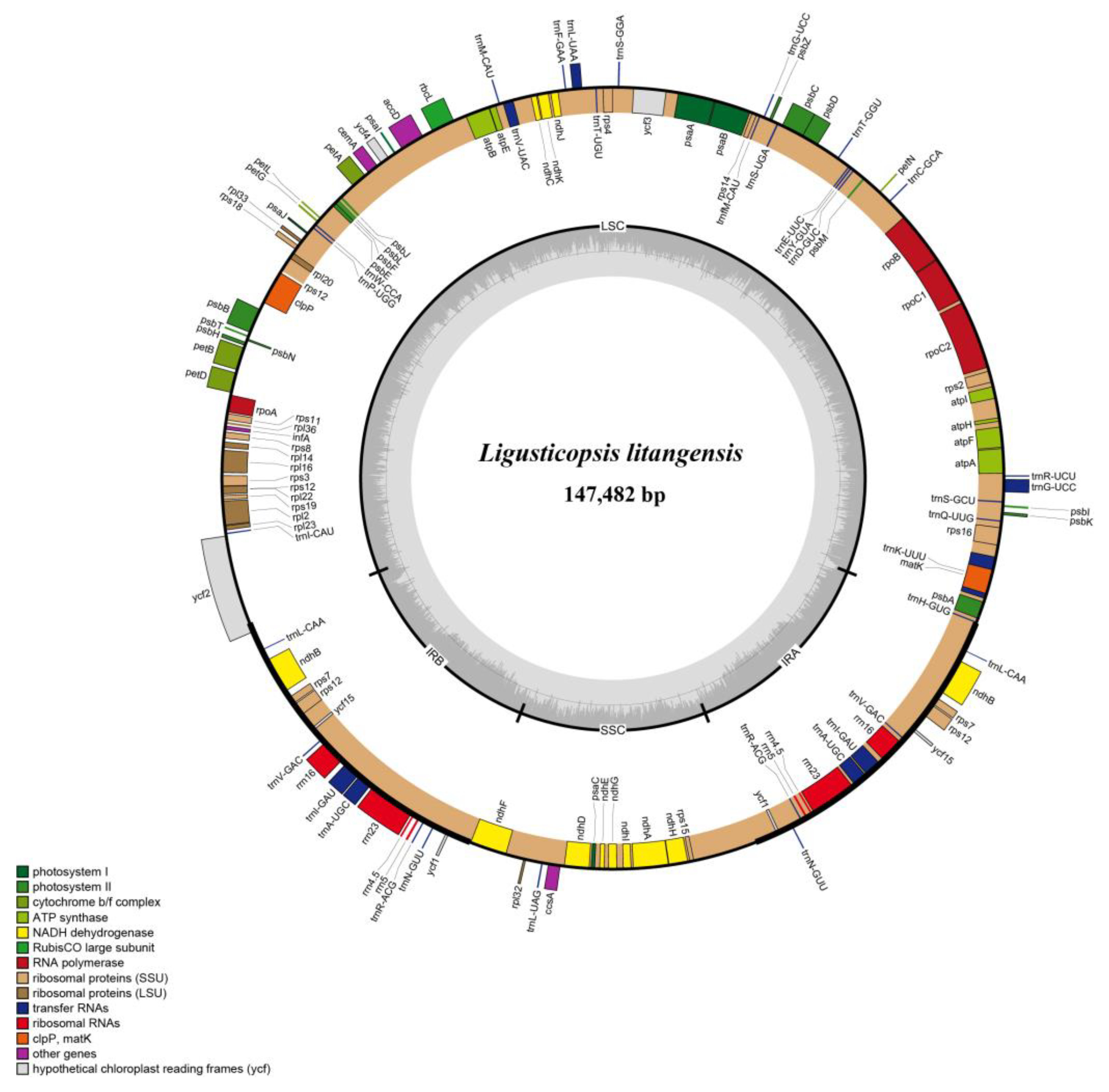
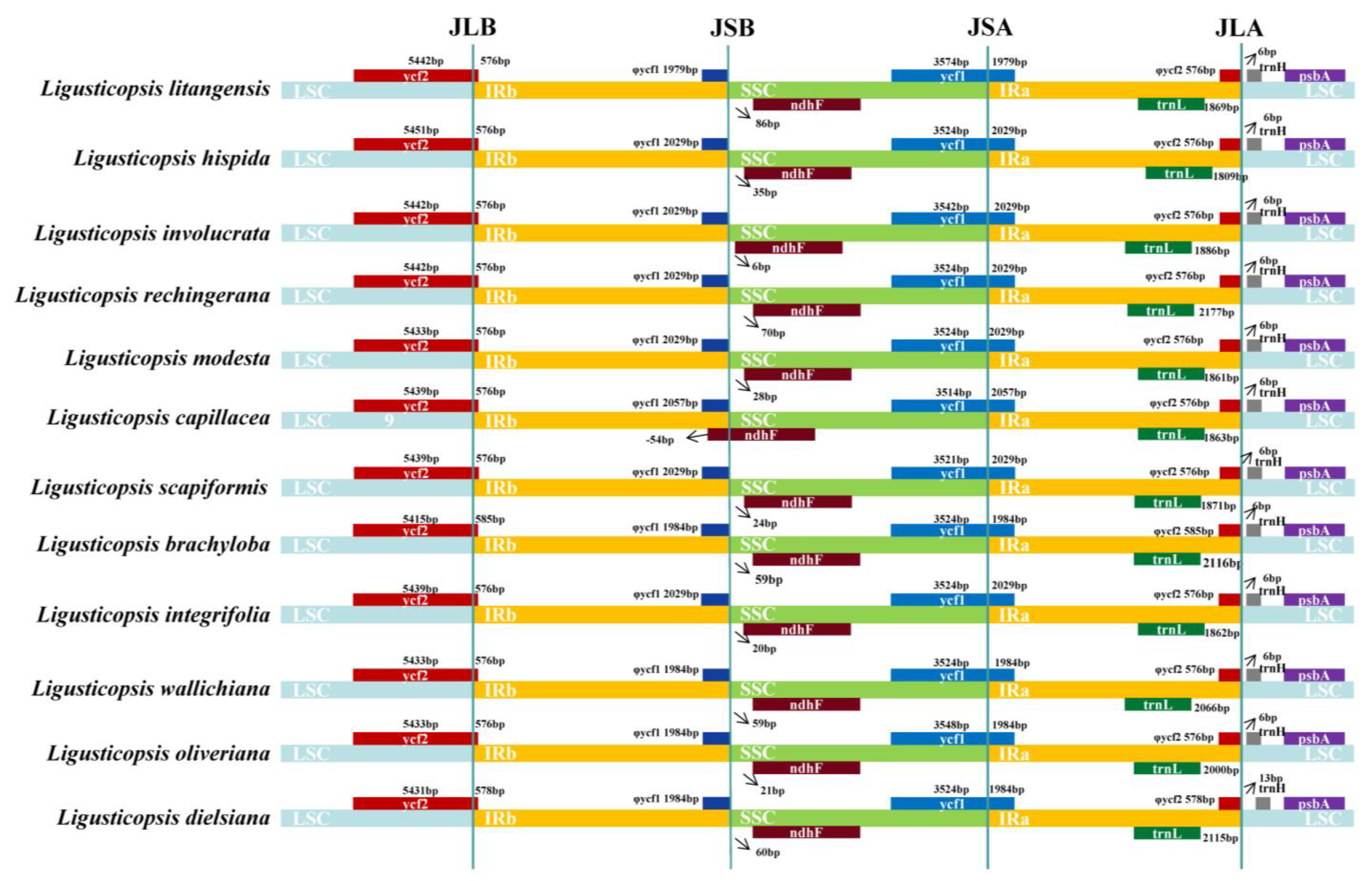
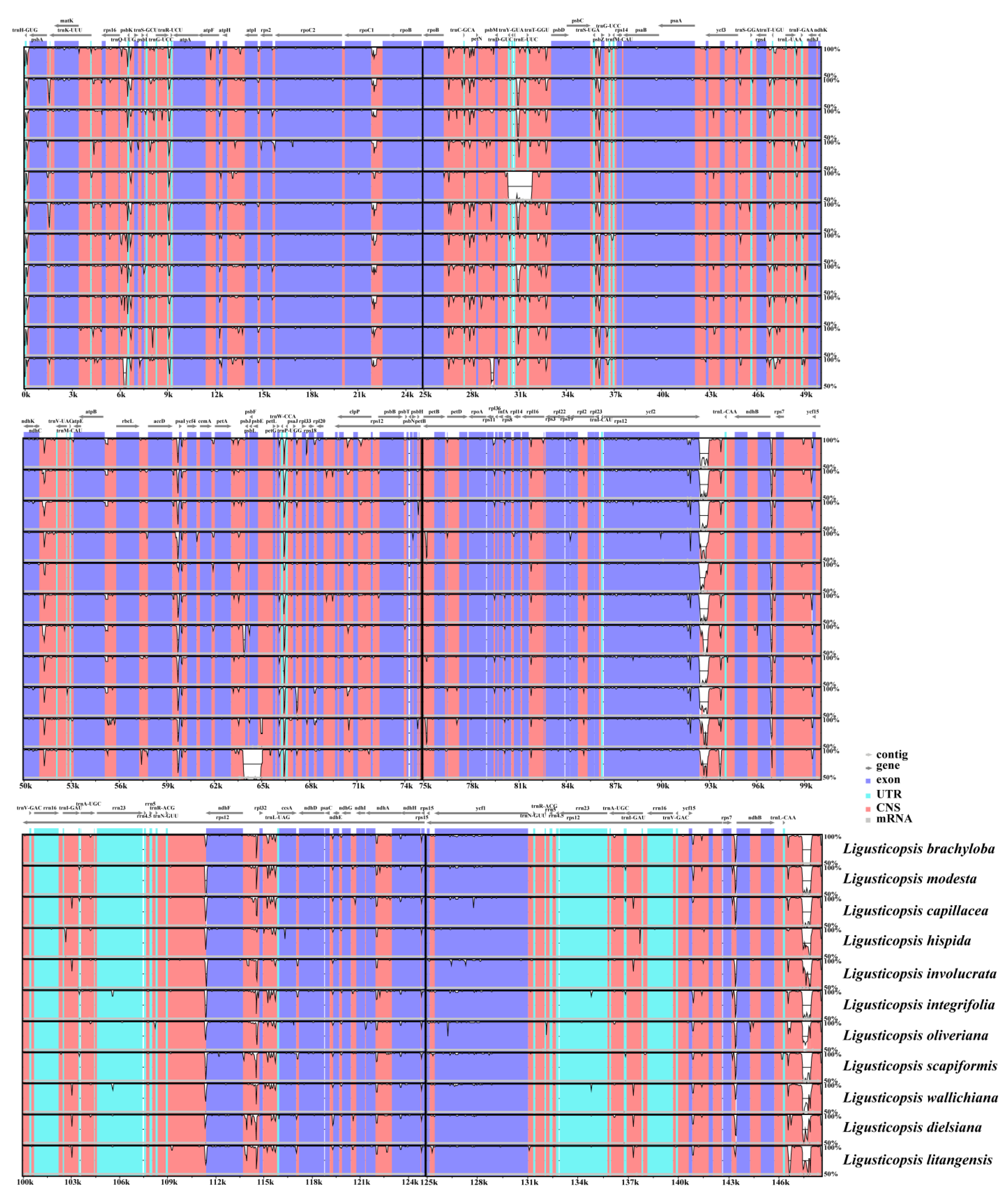
2.2. Comparative Plastome Analyses
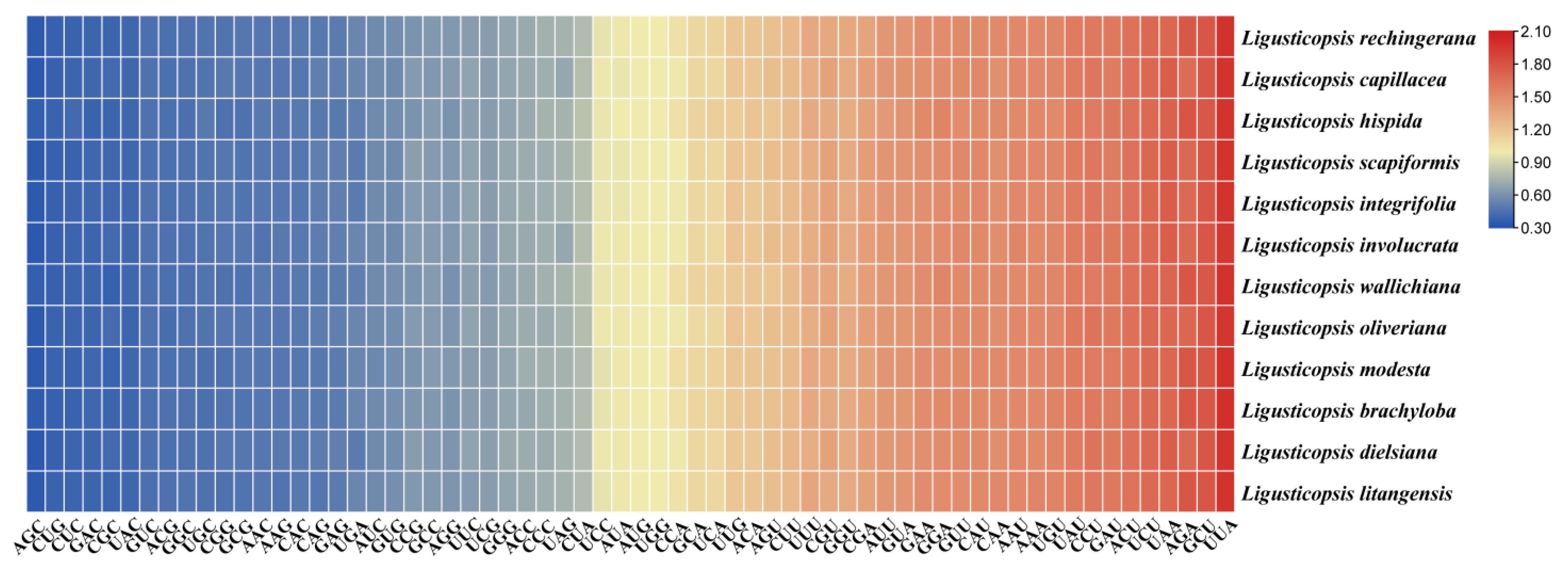
2.3. Codon Usage Analyses
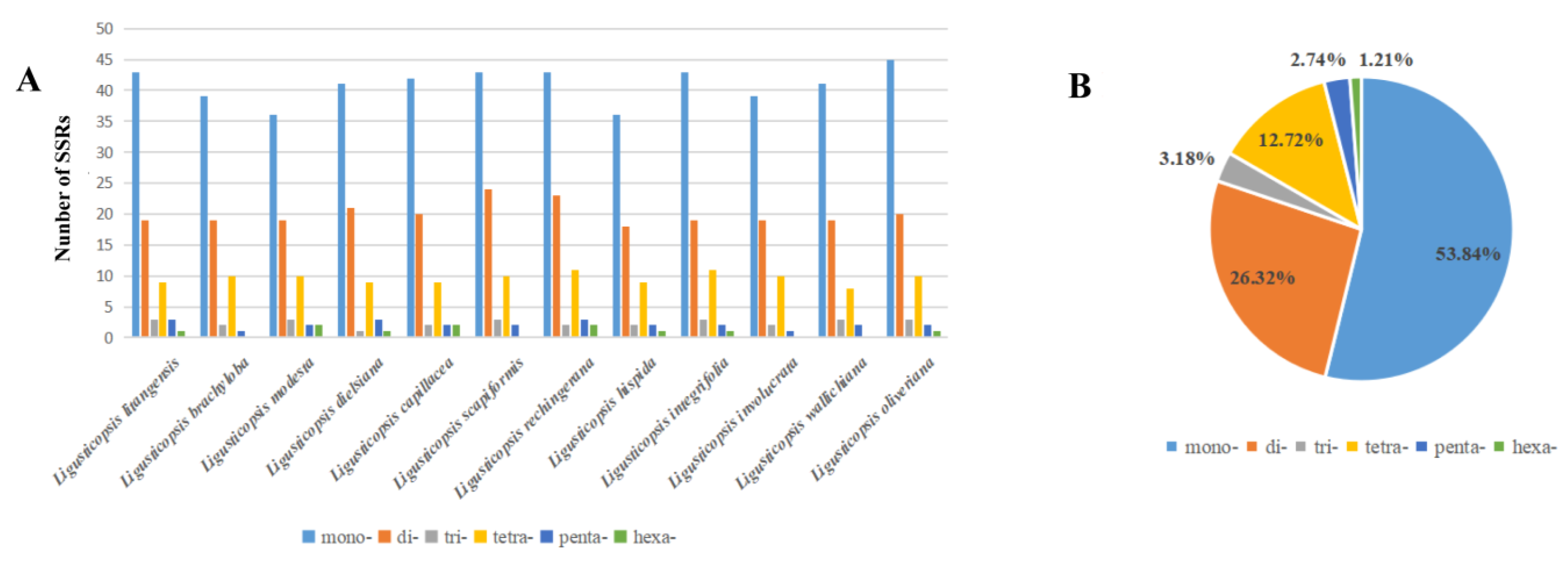
2.4. Simple Sequence Repeats Analyses (SSRs)
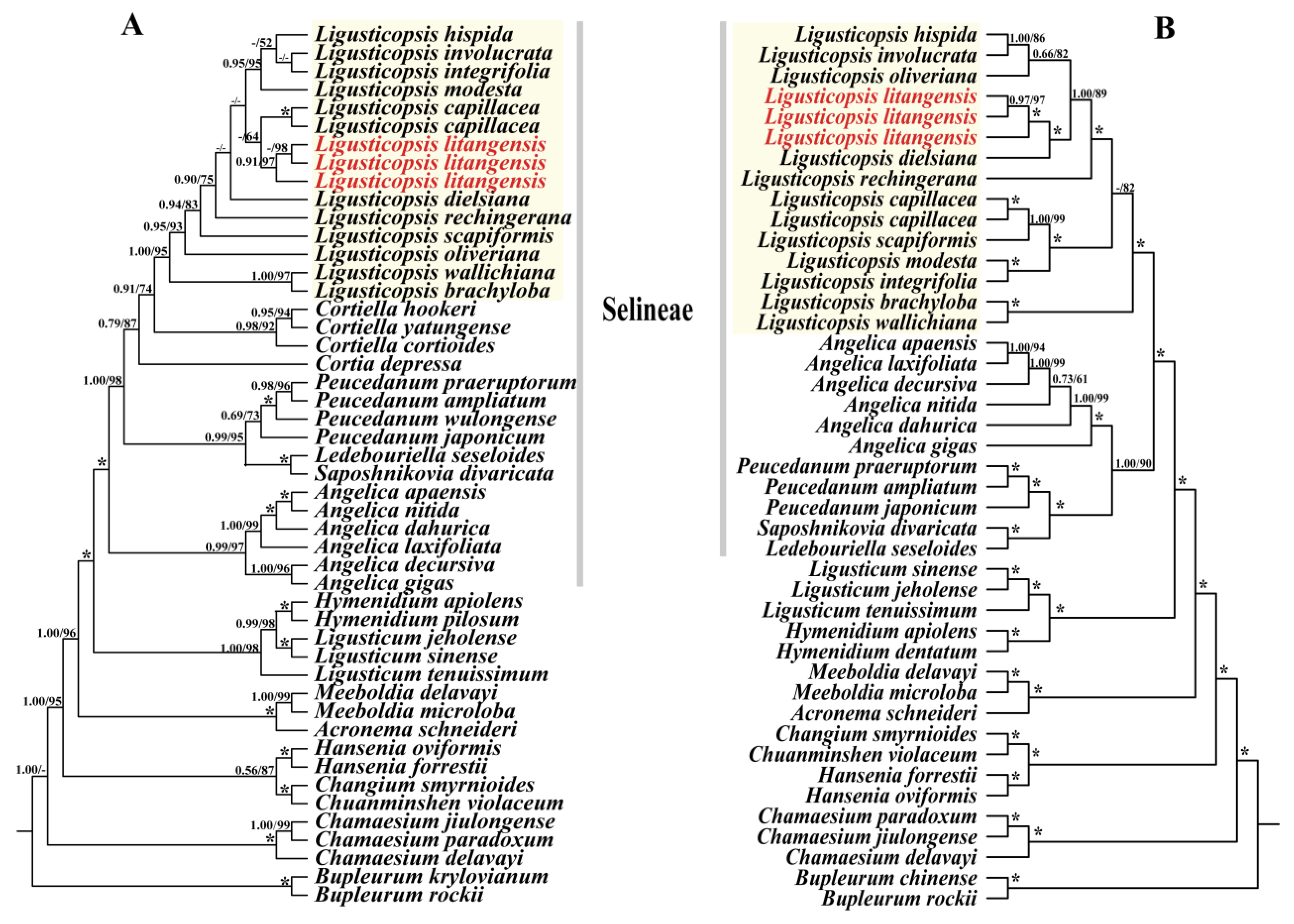
2.5. Phylogenetic Analysis
3. Discussion
3.1. Plastome Characteristic
3.2. Comparison of Ligusticopsis Plastomes
3.3. Phylogenetic Analysis
3.4. Taxonomic Treatment
| 1 | Plants densely villous or strigose | 2 |
| Plants nearly smooth | 4 | |
| 2 | Rays of umbel almost draw from the base, extremely elongated | L. hispida |
| Plants compound umbels with long pedicels, umbrella not elongated | 3 | |
| 3 | Bracts well developed, 2-pinnately divided | L. involucratum |
| Bracts caducous, 1-pinnately divided | L. capillaceum | |
| 4 | Umbels predominantly simple, rays drawn from base very unequal | L. litangensis |
| Compound umbels, rays subequal | 5 | |
| 5 | Basal leaves and lower stem leaves are 1-pinnately compound | L. integrifolia |
| Basal leaves and lower stem leaves are 2–4-pinnately compound | 6 | |
| 6 | Calyx teeth inconspicuous | 7 |
| Calyx teeth prominent | 8 | |
| 7 | Bracteoles margin not membranous | L. modesta |
| Bracteoles with white membranous margin | L. oliverianum | |
| 8 | Stems unbranched, scapiform, cauline leaves usually absent | L. scapiforme |
| Stems simple or branched, cauline leaves present | 9 | |
| 9 | Calyx teeth linear | 10 |
| Calyx teeth subulate | 11 | |
| 10 | Bracteoles with broad white membranous margins | L. wallichiana |
| Bracteoles without white membranous margins | L. brachyloba | |
| 11 | Leaf blade 1-2-pinnate, ultimate segments ovate to oblong-ovate | L. rechingeriana |
| Leaf blade 3-4-pinnate, ultimate segments linear | L. dielsiana |
4. Materials and Methods
4.1. Plant Sample, Morphological Observation and Anatomy
4.2. DNA Extraction, Sequencing, Assembly and Annotation
4.3. Phylogenetic Analyses
4.4. Comparative Plastome Analyses
4.5. Codon Usage and SSRs Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leute, G.H. Untersuchungen über den verwandtschaftskreis der Gattung Ligusticum L. (Umbelliferae) I. Teil. Ann. Des Nat. Mus. Wien 1969, 73, 55–98. [Google Scholar]
- Pimenov, M.G. Updated checklist of Chinese Umbelliferae: Nomenclature, synonymy, typification, distribution. Turczaninowia 2017, 20, 106–239. [Google Scholar]
- Wang, P.L.; Pu, F.T.; Ma, J.S. Pollen morphology of the genus Ligusticum from China and Its systematic significance. J. Syst. Evol. 1991, 29, 235–245. [Google Scholar]
- Sun, N.; He, X.J. Epidermal morphology of Ligusticum (Apiaceae) from China. Ann. Bot. Fenn. 2010, 47, 261–279. [Google Scholar] [CrossRef]
- Li, Z.X.; Guo, X.L.; Price, M.; Zhou, S.D.; He, X.J. Phylogenetic position of Ligusticopsis (Apiaceae, Apioideae): Evidence from molecular data and carpological characters. AoB Plants 2022, 14, plac008. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Li, Z.X.; Xie, D.F. Plastomes of eight Ligusticum species: Characterization, genome evolution, and phylogenetic relationships. BMC Plant Biol. 2020, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Spalik, K.; Katz-Downie, D.S. Major clades within Apiaceae subfamily Apioideae as inferred by phylogenetic analysis of nrDNA ITS sequences. Plant Divers. Evol. 2010, 128, 111–136. [Google Scholar] [CrossRef]
- Hang, S.; Zhang, J.W.; Deng, T.; David, E. Boufford, Origins and evolution of plant diversity in the Hengduan Mountains. China. Plant Divers. 2017, 39, 161–166. [Google Scholar]
- Qiu, Y.X.; Fu, C.X. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenetics Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef]
- Ravi, V.; Khurana, J.P.; Tyagi, A.K.; Khurana, P. An update on chloroplast genomes. Plant Syst. Evol. 2008, 271, 101–122. [Google Scholar] [CrossRef]
- Guisinger, M.M.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Extreme reconfguration of plastid genomes in the angiosperm family Geraniaceae: Rearrangements, repeats, and codon usage. Mol. Biol. Evol. 2011, 28, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Cai, Z.; Raubeson, L.A.; Daniell, H. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifes genome-scale evolutionary patterns. Proc. Natl. Acad. Sci. USA 2007, 104, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Nock, C.J.; Waters, D.L.E.; Edwards, M.A.; Bowen, S.G.; Rice, N.; Cordeiro, G.M. Chloroplast genome sequences from total DNA for plant identifcation. Plant Biotechnol. J. 2011, 9, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.O.; Joh, H.J.; Kim, K.; Lee, S.C.; Kim, N.H.; Park, J.Y. Dynamic chloroplast genome rearrangement and DNA barcoding for three Apiaceae species known as the medicinal herb “Bang-Poong”. Int. J. Mol. Sci. 2019, 20, 2196. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Lim, C.E.; Lee, B.Y. The complete chloroplast genome sequence of Peucedanum chujaense (Apiaceae), an endemic species to Korea. Mitochondrial DNA B Resour. 2020, 5, 1937–1938. [Google Scholar] [CrossRef]
- Mwanzia, V.M.; He, D.X.; Gichira, A.W.; Li, Y.; Ngarega, B.K.; Karichu, M.J. The complete plastome sequences of fve Aponogeton species (Aponogetonaceae): Insights into the structural organization and mutational hotspots. Plant Divers. 2020, 42, 334–342. [Google Scholar] [CrossRef]
- Mehmood, F.; Rahim, A.; Heidari, P.; Ahmed, I.; Peter, P. Comparative plastome analysis of Blumea, with implications for genome evolution and phylogeny of Asteroideae. Eco. Evol. 2021, 11, 7810–7826. [Google Scholar]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Säll, T.; Lind-Halldén, C.; Halldén, C. Evolution of chloroplast mononucleotide microsatellites in Arabidopsis thaliana. Theor. Appl. Genet. 2007, 114, 223. [Google Scholar] [CrossRef]
- Hopkins, M.S.; Casa, A.M.; Wang, T. Discovery and Characterization of Polymorphic Simple Sequence Repeats (SSRs) in Peanut. Crop Sci. 1999, 39, 1243–1247. [Google Scholar] [CrossRef]
- Li, J.; Xie, D.F.; Guo, X.L.; Zheng, Z.Y.; He, X.J.; Zhou, S.D. Comparative analysis of the complete plastid genome of five Bupleurum species and new insights into DNA barcoding and phylogenetic relationship. Plants 2020, 9, 543. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, F.A.; Shahzadi, I.; Waseem, S.; Mirza, B.; Ahmed, I.; Waheed, M.T. Chloroplast genome of Hibiscus rosasinensis (Malvaceae): Comparative analyses and identifcation of mutational hotspots. Genomics 2020, 112, 581–591. [Google Scholar]
- Kim, K.J.; Lee, H.L. Complete chloroplast genome sequences from Korean ginseng (Panax schinseng Nees) and comparative analysis of sequence evolution among 17 vascular plants. DNA Res. 2004, 11, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Menezes, A.P.A.; Resende-Moreira, L.C.; Buzatti, R.S.O.; Nazareno, A.G.; Carlsen, M.; Lobo, F.P.; Kalapothakis, E.; Lovato, M.B. Chloroplast genomes of Byrsonima species (Mal pighiaceae): Comparative analysis and screening of high divergence sequences. Sci. Rep. 2018, 8, 2210. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D.; Nugent, J.M.; Herbon, L.A. Unusual structure of geranium chloroplast DNA: A triplesized inverted repeat, extensive gene duplications, multiple inversions, and two repeat families. Proc. Natl. Acad. Sci. USA 1987, 84, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.K.; Ruhlman, T.A. Plastid Genomes of Seed Plants. In Genomics of Chloroplasts and Mitochondria; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Choi, S.A.; Kim, Y.J.; Kim, K.Y. The complete chloroplast genome sequence of the medicinal plant, Angelica gigas (Apiaceae). Mitochondrial DNA Part B 2016, 1, 280–281. [Google Scholar] [CrossRef]
- Kim, C.K.; Jin, M.W.; Kim, Y.K. The complete mitochondrial genome sequences of Bupleurum falcatum (Apiales: Apiaceae). Mitochondrial DNA Part B 2020, 5, 2576–2577. [Google Scholar] [CrossRef]
- Korpelainen, H. The evolutionary processes of mitochondrial and chloroplast genomes differ from those of nuclear genomes. Sci. Nat. 2004, 91, 505–518. [Google Scholar] [CrossRef]
- Wicke, S.; Schneeweiss, G.M.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef]
- Shaw, J.; Shafer, H.L.; Leonard, O.R.; Kovach, M.J.; Schorr, M.; Morris, A.B. Chloroplast DNA sequence utility for the lowest phylogenetic and phylogeographic inferences in angiosperms, the tortoise and the hare IV. Am. J. Bot. 2014, 101, 1987–2004. [Google Scholar] [CrossRef]
- Tonti-Filippini, J.; Nevill, P.G.; Dixon, K.; Small, I. What can we do with 1000 plastid genomes? Plant J. 2017, 90, e808–e818. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: San Diego, CA, USA, 2019; pp. 315–322. [Google Scholar]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: Denovo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.L.; Jakovlic, I. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Marc, L.; Oliver, D.; Sabine, K.; Ralph, B. OrganellarGenomeDRAW-a suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575. [Google Scholar]
- Wen, J.; Xie, D.F.; Price, M.; Ren, T.; Deng, Y.; Gui, L. Backbone phylogeny and evolution of Apioideae (Apiaceae): New insights from phylogenomic analyses of plastome data. Mol. Phylogenet. Evol. 2021, 161, 107–183. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.Y.; Chen, X.F.; Tan, J.B. The complete chloroplast genome of Ostericum palustre (Apiaceae). Mitochondrial DNA Part B 2020, 5, 1357–1358. [Google Scholar] [CrossRef]
- Xie, D.F.; Yu, H.X.; Price, M. Phylogeny of Chinese Allium species in section Daghestanica and adaptive evolution of Allium (Amaryllidaceae, Allioideae) species revealed by the chloroplast complete genome. Front. Plant Sci. 2019, 10, 460. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.F.; Tan, J.B.; Yu, Y. Insights into phylogeny, age and evolution of Allium (Amaryllidaceae) based on the whole plastome sequences. Ann. Bot. 2020, 125, 1039–1055. [Google Scholar] [CrossRef]
- Xie, F.M.; Xie, D.F.; Xie, C. Adaptation evolution and phylogenetic analyses of species in Chinese Allium section Pallasia and related species based on complete chloroplast genome sequences. BioMed Res. Int. 2020, 2020, 8542797. [Google Scholar] [CrossRef]
- Neuhaus, H.E.; Emes, M.J. Nonphotosynthetic Metabolism in Plastids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 111–140. [Google Scholar] [CrossRef]
- Jansen, R.K.; Raubeson, L.A.; Boore, J.L. Methods for Obtaining and Analyzing Whole Chloroplast Genome Sequences. Methods Enzymol. 2005, 395, 348–384. [Google Scholar]
- Amiryousefi, A.; Hyvonen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Lior, P.; Alexander, P. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273. [Google Scholar] [CrossRef] [PubMed]
- Peden, J.F. Analysis of Codon Usage. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1999. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bryan, G.J.; Mcnicoll, J.; Ramsay, G. Polymorphic simple sequence repeats markers in chloroplast genomes of Solanaceous plants. Theor. Appl. Genet. 1999, 99, 859–867. [Google Scholar] [CrossRef]
- Thiet, T.; Michalek, W.; Varshney, R.K. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | LSC Length (bp) | SSC Length (bp) | IR Length (bp) | Total Genome | Number of Genes | ||||
|---|---|---|---|---|---|---|---|---|---|
| Length (bp) | GC (%) | Total | CDS | rRNA | tRNA | ||||
| L. litangensis | 91,559 | 17,669 | 19,127 | 147,482 | 37.4 | 129 | 85 | 8 | 36 |
| L. dielsiana | 91,666 | 17,582 | 19,415 | 148,078 | 37.4 | 129 | 85 | 8 | 36 |
| L. capillacea | 91,907 | 17,503 | 19,199 | 147,808 | 37.5 | 129 | 85 | 8 | 36 |
| L. scapiformis | 92,214 | 17,581 | 19,156 | 148,107 | 37.5 | 129 | 85 | 8 | 36 |
| L. rechingerana | 91,813 | 17,654 | 19,529 | 148,525 | 37.3 | 129 | 85 | 8 | 36 |
| L. brachyloba | 92,265 | 17,588 | 19,390 | 148,633 | 37.4 | 129 | 85 | 8 | 36 |
| L. hispida | 91,846 | 17,627 | 19,162 | 147,797 | 37.4 | 129 | 85 | 8 | 36 |
| L. integrifolia | 92,305 | 17,575 | 19,158 | 148,196 | 37.5 | 129 | 85 | 8 | 36 |
| L. involucrata | 91,782 | 17,560 | 19,205 | 147,752 | 37.4 | 129 | 85 | 8 | 36 |
| L. modesta | 92,247 | 17,568 | 19,159 | 148,133 | 37.5 | 129 | 85 | 8 | 36 |
| L. wallichiana | 92,281 | 17,567 | 19,373 | 148,594 | 37.4 | 129 | 85 | 8 | 36 |
| L. oliveriana | 92,262 | 17,558 | 19,279 | 148,378 | 37.5 | 129 | 85 | 8 | 36 |
| L. litangensis | 91,559 | 17,669 | 19,127 | 147,482 | 37.4 | 129 | 85 | 8 | 36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, R.; Aou, X.; Song, B.; Li, Z.; He, X.; Zhou, S. Plastid Phylogenomic Analyses Reveal a Cryptic Species of Ligusticopsis (Apiaceae, Angiosperms). Int. J. Mol. Sci. 2023, 24, 7419. https://doi.org/10.3390/ijms24087419
Tian R, Aou X, Song B, Li Z, He X, Zhou S. Plastid Phylogenomic Analyses Reveal a Cryptic Species of Ligusticopsis (Apiaceae, Angiosperms). International Journal of Molecular Sciences. 2023; 24(8):7419. https://doi.org/10.3390/ijms24087419
Chicago/Turabian StyleTian, Rongming, Xueyimu Aou, Boni Song, Zixuan Li, Xingjin He, and Songdong Zhou. 2023. "Plastid Phylogenomic Analyses Reveal a Cryptic Species of Ligusticopsis (Apiaceae, Angiosperms)" International Journal of Molecular Sciences 24, no. 8: 7419. https://doi.org/10.3390/ijms24087419