
Genetic Variability of Tabebuia rosea (Bignoniaceae) from Plantations and Remnant Populations in the Mayan Forest
 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling Strategy
2.3. DNA Isolation, SSR, and ISSR Scoring Analyses
2.4. Data Analyses
2.5. Genetic Structure
2.6. Genetic Differentiation and Genetic Diversity
2.7. Genetic Bottlenecks
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manel, S.; Schwartz, M.K.; Luikart, G.; Taberlet, P. Landscape Genetics: Combining Landscape Ecology and Population Genetics. Trends Ecol. Evol. 2003, 18, 189–197. [Google Scholar] [CrossRef]
- Porter-Bolland, L.; Bonilla-Moheno, M.; Garcia-Frapolli, E.; Morteo-Montiel, S. Forest Ecosystems and Conservation. In Biodiversity and Conservation of the Yucatán Peninsula; Islebe, G., Calmé, S., León-Cortés, J., Schmook, B., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 377–398. [Google Scholar]
- Zizumbo-Villarreal, D.; Colunga-GarcíaMarín, P. Origin of Agriculture and Plant Domestication in West Mesoamerica. Genet. Resour. Crop Evol. 2010, 57, 813–825. [Google Scholar] [CrossRef]
- Turner, B.; Villar, S.C.; Foster, D.; Geoghegan, J.; Keys, E.; Klepeis, P.; Lawrence, D.; Mendoza, P.M.; Manson, S.; Ogneva-Himmelberger, Y.; et al. Deforestation in the Southern Yucatán Peninsular Region: An Integrative Approach. For. Ecol. Manag. 2001, 154, 353–370. [Google Scholar] [CrossRef]
- Oglesby, R.J.; Sever, T.L.; Saturno, W.; Erickson, D.J.; Srikishen, J. Collapse of the Maya: Could Deforestation Have Contributed? J. Geophys. Res. 2010, 115, D12106. [Google Scholar] [CrossRef]
- Ng, K.K.S.; Lee, S.L.; Ueno, S. Impact of Selective Logging on Genetic Diversity of Two Tropical Tree Species with Contrasting Breeding Systems Using Direct Comparison and Simulation Methods. For. Ecol. Manag. 2009, 257, 107–116. [Google Scholar] [CrossRef]
- Carneiro, F.S.; Lacerda, A.E.B.; Lemes, M.R.; Gribel, R.; Kanashiro, M.; Wadt, L.H.O.; Sebbenn, A.M. Effects of Selective Logging on the Mating System and Pollen Dispersal of Hymenaea courbaril L. (Leguminosae) in the Eastern Brazilian Amazon as Revealed by Microsatellite Analysis. For. Ecol. Manag. 2011, 262, 1758–1765. [Google Scholar] [CrossRef]
- Cascante, A.; Quesada, M.; Lobo, J.J.; Fuchs, E.A. Effects of Dry Tropical Forest Fragmentation on the Reproductive Success and Genetic Structure of the Tree Samanea Saman. Conserv. Biol. 2002, 16, 137–147. [Google Scholar] [CrossRef]
- Duminil, J.; Hardy, O.J.; Petit, R.J. Plant Traits Correlated with Generation Time Directly Affect Inbreeding Depression and Mating System and Indirectly Genetic Structure. BMC Evol. Biol. 2009, 9, 177. [Google Scholar] [CrossRef]
- Kitzmiller, J.H. Managing Genetic Diversity in a Tree Improvement Program. For. Ecol. Manag. 1990, 35, 131–149. [Google Scholar] [CrossRef]
- Chwedorzewska, K.J.; Galera, H.; Kosiński, I. Plantations of Convallaria majalis L. as a Threat to the Natural Stands of the Species: Genetic Variability of the Cultivated Plants and Natural Populations. Biol. Conserv. 2008, 141, 2619–2624. [Google Scholar] [CrossRef]
- Solís-Guillén, I.; Chaires-Pacheco, M.; Juárez-Gómez, J.; O’Connor-Sánchez, A.; Peña-Ramírez, Y.J. Development of an Ideotype-Based Selection Tool for Native Tropical Tree Breeding by Smallholder Planters in Mexico’s Maya Forest. Small Scale For. 2017, 16, 521–534. [Google Scholar] [CrossRef]
- Jordan, R.; Breed, M.F.; Prober, S.M.; Miller, A.D.; Hoffmann, A.A. How Well Do Revegetation Plantings Capture Genetic Diversity? Biol. Lett. 2019, 15, 20190460. [Google Scholar] [CrossRef] [PubMed]
- İçgen, Y.; Kaya, Z.; Çengel, B.; Velioğlu, E.; Öztürk, H.; Önde, S. Potential Impact of Forest Management and Tree Improvement on Genetic Diversity of Turkish Red Pine (Pinus brutia Ten.) Plantations in Turkey. For. Ecol. Manag. 2006, 225, 328–336. [Google Scholar] [CrossRef]
- Vander Mijnsbrugge, K.; Bischoff, A.; Smith, B. A Question of Origin: Where and How to Collect Seed for Ecological Restoration. Basic Appl. Ecol. 2010, 11, 300–311. [Google Scholar] [CrossRef]
- Henery, M.L. The Constraints of Selecting for Insect Resistance in Plantation Trees. Agric. For. Entomol. 2011, 13, 111–120. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors Influencing Levels of Genetic Diversity in Woody Plant Species. In Population Genetics of Forest Trees; Adams, W.T., Strauss, S.H., Copes, D.L., Griffin, A.R., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 95–124. ISBN 978-94-011-2815-5. [Google Scholar]
- Hamrick, J.; Godt, M. Effects of Life History Traits on Genetic Diversity in Plant Species. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for Evolutionary Responses to Climate Change—Evidence from Tree Populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef]
- Hackenberg, D.; Twell, D. The Evolution and Patterning of Male Gametophyte Development. Curr. Top. Dev. Biol. 2019, 131, 257–298. [Google Scholar]
- de Candolle, A.P. Prodromus Systematis Naturalis Regni Vegetabilis, Sive, Enumeratio Contracta Ordinum generum Specierumque Plantarum Huc Usque Cognitarium, Juxta Methodi Naturalis, Normas Digesta; Treuttel et Würtz: London, UK, 1864; Volume 15. [Google Scholar]
- Gentry, A.H. A Synopsis of Bignoniaceae Ethnobotany and Economic Botany. Ann. Missouri Bot. Gard. 1992, 79, 53. [Google Scholar] [CrossRef]
- Comisión Nacional Forestal. Estado Que Guarda el Sector Forestal en México 2021, 1st ed.; Ladrón de Guevara Alafita, E., Venegas López, M., Jaimes Reynada, Z., León Corrales, M.Á., Escoto Prado, J.C., Eds.; CONAFOR: Zapopan, Mexico, 2022. [Google Scholar]
- Villalobos-Zapata, G.J.; Mendoza Vega, J. La Biodiversidad en Campeche: Estudio de Estado; CONABIO, Gobierno del Estado de Campeche, UAC, ECOSUR: Campeche, Mexico, 2010; ISBN 9786077887225. [Google Scholar]
- Castañeda-Navarrete, J. Homegarden Diversity and Food Security in Southern Mexico. Food Secur. 2021, 13, 669–683. [Google Scholar] [CrossRef]
- Pimenta, J.M.A.; Felix, F.C.; de Araújo, J.S.O.; Fajardo, C.G.; Pacheco, M.V. Selection of ISSR Molecular Primers for Studies of Genetic Diversity in Handroanthus impetiginosus (Mart. Ex Dc.) Mattos. Rev. Caatinga 2022, 35, 231–238. [Google Scholar] [CrossRef]
- Vanden Broeck, A.; Villar, M.; Van Bockstaele, E.; VanSlycken, J. Natural Hybridization between Cultivated Poplars and Their Wild Relatives: Evidence and Consequences for Native Poplar Populations. Ann. For. Sci. 2005, 62, 601–613. [Google Scholar] [CrossRef]
- Moreira, P.A.; Fernandes, G.W.; Collevatti, R.G. Fragmentation and Spatial Genetic Structure in Tabebuia ochracea (Bignoniaceae) a Seasonally Dry Neotropical Tree. For. Ecol. Manag. 2009, 258, 2690–2695. [Google Scholar] [CrossRef]
- Braga, A.C.; Reis, A.M.M.; Leoi, L.T.; Pereira, R.W.; CollevattiI, R.G. Development and Characterization of Microsatellite Markers for the Tropical Tree Species Tabebuia aurea (Bignoniaceae). Mol. Ecol. Notes 2006, 7, 53–56. [Google Scholar] [CrossRef]
- López, A.M.; Marulanda, M.L.; Ospina, C.M. Characterizing Tabebuia rosea (Bertol.) DC. Using Microsatellites in Provenance and Progeny Trials in Colombia. In Molecular Approaches to Genetic Diversity; Caliskan, M., Oz, G.C., Kavakli, I.H., Ozcan, B., Eds.; InTech: London, UK, 2015; pp. 1–18. ISBN 978-953-51-2042-1. [Google Scholar]
- Feres, J.M.; Martinez, M.L.L.; Martinez, A.A.; Mestriner, M.A.; Alzate-Marin, A.L. Transferability and Characterization of Nine Microsatellite Markers for the Tropical Tree Species Tabebuia roseo-alba. Mol. Ecol. Resour. 2009, 9, 434–437. [Google Scholar] [CrossRef]
- Comisión Nacional para el Comocimiento y uso de la Biodiversidad CONABIO Proyecto Corredor Biológico Mesoamericano—México. Available online: https://www.biodiversidad.gob.mx/region/cbmm (accessed on 2 August 2023).
- Vinson, C.C.; Mangaravite, E.; Sebbenn, A.M.; Lander, T.A. Using Molecular Markers to Investigate Genetic Diversity, Mating System and Gene Flow of Neotropical Trees. Rev. Bras. Bot. 2018, 41, 481–496. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Dai, S.-L. Genetic Variation within and among Populations of Orychophragmus violaceus (Cruciferae) in China as Detected by ISSR Analysis. Genet. Resour. Crop Evol. 2010, 57, 55–64. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística Geografía e Informática INEGI Fisiografía de México. Available online: https://www.inegi.org.mx/temas/fisiografia/#Mapa (accessed on 2 August 2023).
- Secretaría de Medio Ambiente y Recursos Naturales SEMARNAT Atlas Digital Geográfico de México. Available online: http://gisviewer.semarnat.gob.mx/aplicaciones/Atlas2015/index.html (accessed on 12 December 2021).
- Van Oosterhout, C.; Hutchinson, W.F.; Willis, D.P.M.; Shipley, P. Micro-Checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Chybicki, I.J.; Burczyk, J. Simultaneous Estimation of Null Alleles and Inbreeding Coefficients. J. Hered. 2009, 100, 106–113. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Corander, J.; Sirén, J.; Arjas, E. Bayesian Spatial Modeling of Genetic Population Structure. Comput. Stat. 2008, 23, 111–129. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A Cluster Matching and Permutation Program for Dealing with Label Switching and Multimodality in Analysis of Population Structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, N.A. Distruct: A Program for the Graphical Display of Population Structure. Mol. Ecol. Notes 2003, 4, 137–138. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Lewis, P.O.; Dey, D.K. A Bayesian Approach to Inferring Population Structure from Dominant Markers. Mol. Ecol. 2002, 11, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Milligan, B.G. Analysis of Population Genetic Structure with RAPD Markers. Mol. Ecol. 1994, 3, 91–99. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Lewis, P.O. Hickory: A Package for Analysis of Population Genetic Data, Version 1.1; Free Software Foundation: Boston, MA, USA, 2007.
- Vekemans, X. AFLP-SURV. 2002. Available online: https://ebe.ulb.ac.be/ebe/AFLP-SURV.html (accessed on 1 February 2022).
- Felsenstein, J. PHYLIP. 2021. Available online: https://evolution.genetics.washington.edu/phylip.html (accessed on 1 February 2022).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A. Estimating Population Structure in Diploids with Multilocus Dominant DNA Markers. Mol. Ecol. 1999, 8, 907–913. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and Power Analysis of Two Tests for Detecting Recent Population Bottlenecks from Allele Frequency Data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Young, A.; Boyle, T.; Brown, T. The Population Genetic Consequences of Habitat Fragmentation for Plants. Trends Ecol. Evol. 1996, 11, 413–418. [Google Scholar] [CrossRef]
- Feres, J.M.; Sebbenn, A.M.; Guidugli, M.C.; Mestriner, M.A.; Moraes, M.L.T.; Alzate-Marin, A.L. Mating System Parameters at Hierarchical Levels of Fruits, Individuals and Populations in the Brazilian Insect-Pollinated Tropical Tree, Tabebuia roseo-alba (Bignoniaceae). Conserv. Genet. 2012, 13, 393–405. [Google Scholar] [CrossRef]
- Stacy, E.A.; Hamrick, J.L.; Nason, J.D.; Hubbell, S.P.; Foster, R.B.; Condit, R. Pollen Dispersal in Low-Density Populations of Three Neotropical Tree Species. Am. Nat. 1996, 148, 275–298. [Google Scholar] [CrossRef]
- Dick, C.W.; Hardy, O.J.; Jones, F.A.; Petit, R.J. Spatial Scales of Pollen and Seed-Mediated Gene Flow in Tropical Rain Forest Trees. Trop. Plant Biol. 2008, 1, 20–33. [Google Scholar] [CrossRef]
- Stefenon, V.M.; Gailing, O.; Finkeldey, R. Genetic Structure of Plantations and the Conservation of Genetic Resources of Brazilian Pine (Araucaria Angustifolia). For. Ecol. Manage. 2008, 255, 2718–2725. [Google Scholar] [CrossRef]
- Ferreira, D.K.; Nazareno, A.G.; Mantovani, A.; Bittencourt, R.; Sebbenn, A.M.; dos Reis, M.S. Genetic Analysis of 50-Year Old Brazilian Pine (Araucaria angustifolia) Plantations: Implications for Conservation Planning. Conserv. Genet. 2012, 13, 435–442. [Google Scholar] [CrossRef]
- Collevatti, R.G.; Terribile, L.C.; Rabelo, S.G.; Lima-Ribeiro, M.S. Relaxed Random Walk Model Coupled with Ecological Niche Modeling Unravel the Dispersal Dynamics of a Neotropical Savanna Tree Species in the Deeper Quaternary. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Tapia-Gómez, C.A.; Estrada-Medina, H.; del Ruenes-Morales, M.; Montañez-Escalante, P.I.; Jiménez-Osornio, J.J. Growing Out of the Tropical Forests: Gene Flow of Native Mesoamerican Trees Among Forest and Mayan Homegardens. Front. Ecol. Evol. 2021, 9, 1–14. [Google Scholar] [CrossRef]
- Petit, R.J.; Hampe, A. Some Evolutionary Consequences of Being a Tree. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 187–214. [Google Scholar] [CrossRef]
- Sork, V.L.; Smouse, P.E. Genetic Analysis of Landscape Connectivity in Tree Populations. Landsc. Ecol. 2006, 21, 821–836. [Google Scholar] [CrossRef]
- White, G.M.; Boshier, D.H.; Powell, W. Increased Pollen Flow Counteracts Fragmentation in a Tropical Dry Forest: An Example from Swietenia Humilis Zuccarini. Proc. Natl. Acad. Sci. USA 2002, 99, 2038–2042. [Google Scholar] [CrossRef]
- Fageria, M.S.; Rajora, O.P. Effects of Harvesting of Increasing Intensities on Genetic Diversity and Population Structure of White Spruce. Evol. Appl. 2013, 6, 778–794. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
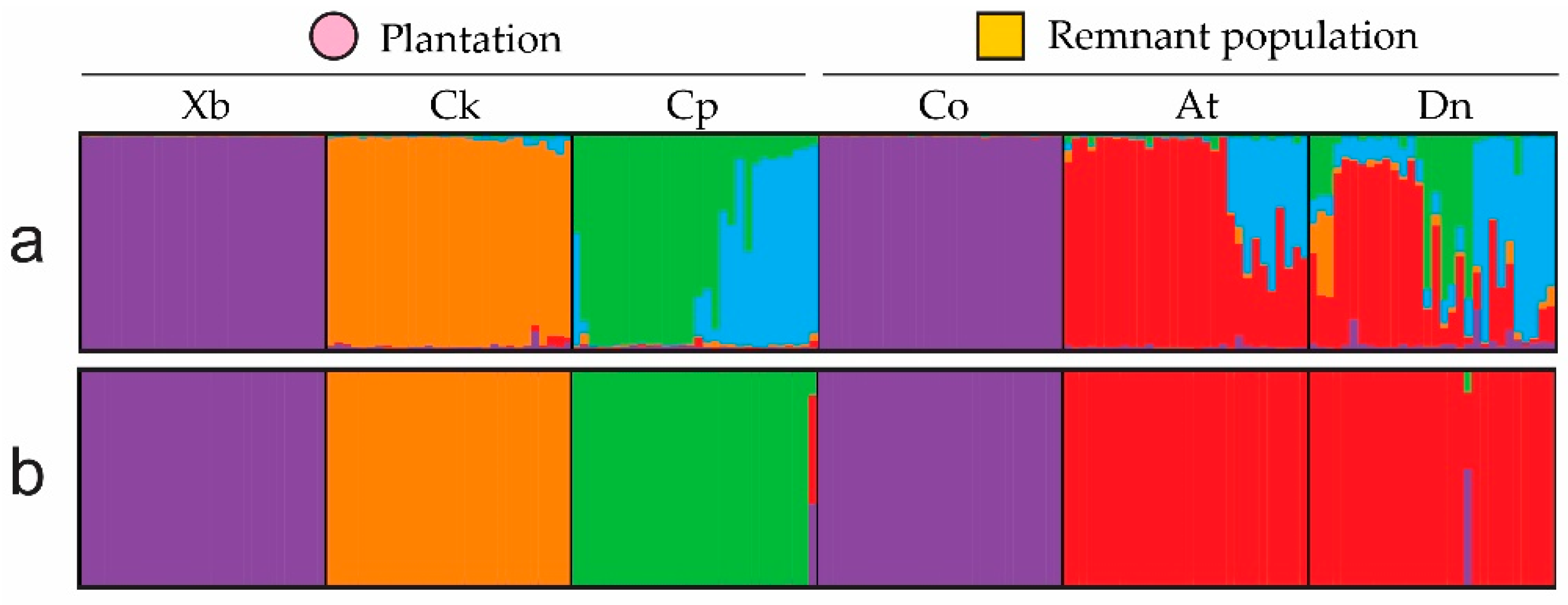{kind=link}
{kind=link}
| Location | Type | Patch Area (ha) | Sampled Area (m2) | Estimated Individuals per ha | Average Distance between Sampled Individuals (m) |
|---|---|---|---|---|---|
| ● Ck | Plantation | 6.0 | 13,780 | 23.22 | 23.76 ± 15.46 |
| ● Cp | Plantation | 0.5 | 49,200 | 65.04 | 17.59 ± 15.29 |
| ● Xb | Plantation | 120.0 | 120,000 | 71.11 | 69.91 ± 51.41 |
| ■ Dn | Remnant | 269.79 | 332,760 | 0.96 | 286.58 ± 409.87 |
| ■ At | Remnant | 66.80 | 104,760 | 3.05 | 92.44 ± 110.48 |
| ■ Co | Remnant | 34.92 | 113,080 | 2.83 | 94.24 ± 133.45 |
| Grouping | n | NG | NA | Ho (SE.) | He (SE.) | F (95% CI) | NPL | PPL | He (SE.) |
|---|---|---|---|---|---|---|---|---|---|
| (a) Population | |||||||||
| ● Ck | 30 | 26 | 15 | 0.760 b (0.133) | 0.840 b (0.133) | 0.08 (0.002–0.20) | 114 | 38.5 | 0.105 b (0.008) |
| ● Cp | 30 | 23 | 16 | 0.774 b (0.156) | 0.838 b (0.150) | 0.06 (0.001–0.18) | 146 | 49.3 | 0.131 b (0.007) |
| ● Xb | 30 | 23 | 3 | 0.416 a (0.234) | 0.521 a (0.212) | 0.07 (0.002–0.18) | 26 | 8.8 | 0.024 a (0.005) |
| ■ Dn | 30 | 25 | 16 | 0.804 b (0.169) | 0.854 b (0.168) | 0.06 (0.001–0.16) | 126 | 42.6 | 0.106 b (0.007) |
| ■ At | 30 | 26 | 12 | 0.752 b (0.166) | 0.802 b (0.121) | 0.03 (0.000–0.10) | 129 | 43.6 | 0.118 b (0.008) |
| ■ Co | 30 | 24 | 3 | 0.418 a (0.232) | 0.474 a (0.251) | 0.06 (0.002–0.16) | 113 | 38.2 | 0.036 a (0.006) |
| (b) Population type | |||||||||
| ● Plantation | 90 | 89 | 23 | 0.451 (0.149) | 0.880 (0.084) | 0.47 (0.435–0.51) | 38 | 12.8 | 0.093 (0.005) |
| ■ Remnant | 90 | 76 | 22 | 0.787 (0.093) | 0.860 (0.100) | 0.06 (0.003–0.18) | 117 | 39.5 | 0.092 (0.006) |
| (c) Clusters | |||||||||
| Cluster 1 | 60 | 59 | 4 | 0.374 c (0.141) | 0.552 c (0.196) | 0.27 (0.055–0.39) | 39 | 13.2 | 0.031 c (0.005) |
| Cluster 2 | 60 | 60 | 20 | 0.594 d (0.206) | 0.849 d (0.142) | 0.28 (0.192–0.34) | 131 | 44.3 | 0.115 d (0.007) |
| Cluster 3 | 30 | 26 | 15 | 0.764 d (0.133) | 0.840 d (0.133) | 0.08 (0.002–0.20) | 114 | 38.5 | 0.105 d (0.008) |
| Cluster 4 | 30 | 23 | 16 | 0.774 d (0.156) | 0.838 d (0.150) | 0.06 (0.001–0.18) | 146 | 49.3 | 0.131 d (0.007) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-González, H.; Raggio, M.; Labrín-Sotomayor, N.Y.; Ferrer, M.M.; Peña-Ramírez, Y.J. Genetic Variability of Tabebuia rosea (Bignoniaceae) from Plantations and Remnant Populations in the Mayan Forest. Forests 2023, 14, 2006. https://doi.org/10.3390/f14102006
Ruiz-González H, Raggio M, Labrín-Sotomayor NY, Ferrer MM, Peña-Ramírez YJ. Genetic Variability of Tabebuia rosea (Bignoniaceae) from Plantations and Remnant Populations in the Mayan Forest. Forests. 2023; 14(10):2006. https://doi.org/10.3390/f14102006
Chicago/Turabian StyleRuiz-González, Hugo, María Raggio, Natalia Y. Labrín-Sotomayor, Miriam M. Ferrer, and Yuri J. Peña-Ramírez. 2023. "Genetic Variability of Tabebuia rosea (Bignoniaceae) from Plantations and Remnant Populations in the Mayan Forest" Forests 14, no. 10: 2006. https://doi.org/10.3390/f14102006