
Softwood Anatomy: A Review
by
 , , , ,
, , , ,
Luis G. Esteban
1,*,
Paloma de Palacios
1,
Immo Heinz
2,
Peter Gasson
3,
Alberto García-Iruela
4 and
Francisco García-Fernández
1 1
Departamento de Sistemas y Recursos Naturales, Escuela Técnica Superior de Ingeniería de Montes, Forestal y del Medio Natural, Universidad Politécnica de Madrid, Ciudad Universitaria, 28040 Madrid, Spain
2
Johann Heinrich von Thünen Institute (TI), Institute of Wood Research, D-21031 Hamburg, Germany
3
Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK
4
Departamento de Ingeniería y Gestión Forestal y Ambiental, Escuela Técnica Superior de Ingeniería de Montes, Forestal y del Medio Natural, Universidad Politécnica de Madrid, Ciudad Universitaria, 28040 Madrid, Spain
*
Author to whom correspondence should be addressed.
Forests 2023, 14(2), 323; https://doi.org/10.3390/f14020323
Submission received: 2 January 2023
/
Revised: 24 January 2023
/
Accepted: 1 February 2023
/
Published: 6 February 2023
(This article belongs to the Special Issue Reviews on Structure and Physical and Mechanical Properties of Wood)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Although conifers have very homogeneous wood anatomical structure, some of their features allow differentiation of genera or groups of genera. Softwood anatomy is reviewed and the authors have added contributions, grouping features in five blocks (axial tracheids, axial parenchyma, rays, resin canals and mineral inclusions), with special attention to their diagnostic value at the family, genus or species levels. The discussion addresses, in axial tracheids: growth rings, intercellular spaces, pit arrangement, presence of scalloped tori, torus extensions, notches, warty layer, organic deposits, helical and callitroid thickenings, bars of Sanio and trabeculae; in axial parenchyma: presence or absence, arrangement and type of transverse end walls; in rays: composition, type of horizontal and end walls and cross-field pitting; in resin canals: epithelium and the presence of normal or traumatic resin canals; and in mineral inclusions: presence or absence. Wood structure is characterised at the family level, discussing the main elements that can be used for diagnosis at the genus level or in groups of genera.
Keywords:
conifers; Araucariaceae; Cupressaceae; gymnosperms; Pinaceae; Podocarpaceae; Sciadopityaceae; softwoods; Taxaceae; wood anatomy1. Gymnosperms
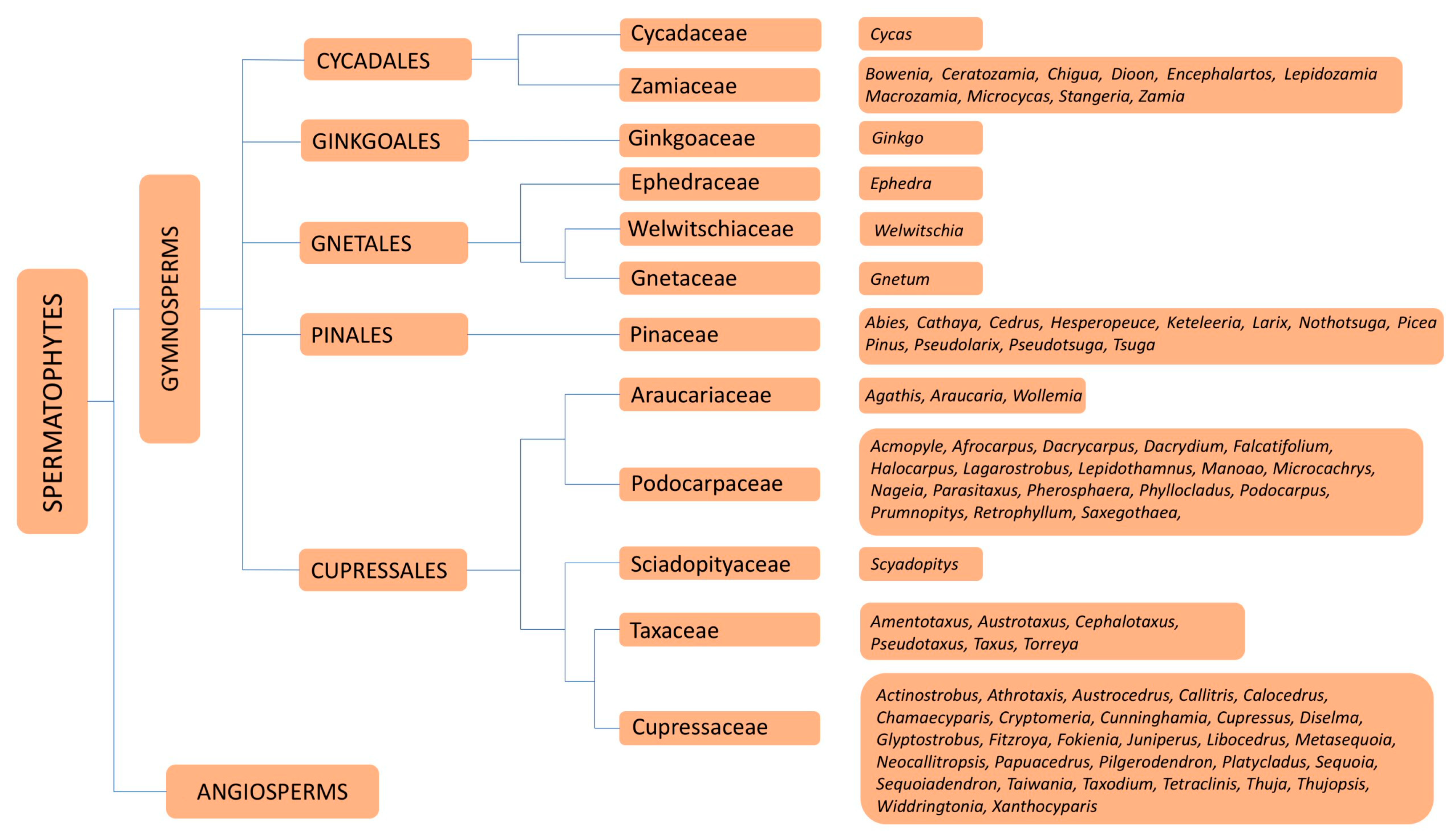
Based on molecular phylogeny, Cole et al. [1] divided the gymnosperms into five orders (Cycadales, Ginkgoales, Gnetales, Pinales and Cupressales) (Figure 1). A total of 669 species, comprising 72 genera in six families (Araucariaceae, Cupressaceae, Pinaceae, Podocarpaceae, Sciadopityaceae and Taxaceae) [2] included in the orders Pinales (Pinaceae) and Cupressales (Araucariaceae, Cupressaceae, Podocarpaceae, Sciadopityaceae and Taxaceae), are known generically by the term conifers (Figure 2) and produce wood in the strict sense (i.e., secondary xylem). The family with the highest number of species is Pinaceae (235), followed by Podocarpaceae (191) and Cupressaceae (170). The most numerous genera by far are Pinus (118) and Podocarpus (115).
The wood produced in Cycadales (Cycadaceae and Zamiaceae) and Gnetales (Ephedraceae, Welwitschiaceae and Gnetaceae) (Figure 3 and Figure 4) differs from conifer wood.
In Ginkgoales, Ginkgo is very similar to Cycas in embryology, but in macromorphology and vegetative anatomy it is more similar to the conifers, suggesting that it may occupy an intermediate phylogenetic position between cycads and conifers [3]. It has secondary growth and produces wood similar to conifers, with abundant druse crystal inclusions in axial parenchyma [4]. Because of its similarity to conifer wood, Ginkgo was included in the IAWA list of microscopic features for softwood identification [5] (Figure 5).
In the genus Cycas, Terrazas [6] confirmed the presence of a first vascular cambium simultaneously active alongside later successive cambia. The secondary xylem of Cycas can be described schematically as the presence of rows of tracheids mixed with thin-walled parenchyma cells.
The nine genera of Zamiaceae (Bowenia Hook., Ceratozamia Brongn., Dioon Lindl., Encephalartos Lehm., Lepidozamia Regel, Macrozamia Miq., Microcycas (Miq.) A.DC., Stangeria T.Moore, Zamia L.) have a similar structure to Cycadaceae (monoxylic), with rows of tracheids and abundant parenchyma cells, generally with high starch content.
The genus Ephedra, the only one in Ephedraceae, develops wood with features characteristic of conifers and Dicotyledonous Angiosperms, causing Carlquist [7] to question which group it belongs to. Some authors [8,9] considered it to clearly be a Gymnosperm, while others [10] described it as an Angiosperm due to the presence of vessels with scalariform perforation plates. The presence of elements characteristic of both large groups (Angiosperm vessels and conifer tracheids) certainly makes this genus very special from an anatomical and evolutionary perspective (Figure 5).
Welwitschia mirabilis Hook. f. produces in its lifetime only two leaves, whose tips necrose as the plant grows. The plant develops a small trunk, where successive cambia develop, producing xylem and phloem, although not annually. It produces vessel elements and tracheids that are always very narrow, and therefore, all secondary xylem can be considered latewood in character [11,12]. Structurally, it is very different from typical conifer or dicotyledonous angiosperm wood, but its individual elements are characteristic of wood (vessels with simple perforation plates, tracheids, small portions of diffuse axial parenchyma between vessels and tracheids of the secondary xylem and rays with exclusively upright cells).
Gnetum also contains tracheids and vessels with secondary growth similar to Ephedra, in tree or bush species, while lianoid species have anomalous cambial growth [13]. The primary cambium quickly ceases activity and successive cambia appear from the outermost secondary phloem towards the inside (Figure 5).
The use of molecular phylogeny continues in the production of numerous changes in conifer systematics and taxonomy. However, this does not affect the study of their anatomy unless the aim is to make an association between anatomy and phylogeny, a good way to answer certain questions, such as the place of Cedrus in Abietoideae. Unlike the other Abietoideae genera, Cedrus is the only genus of this subfamily capable of forming traumatic axial and radial resin canals [14]. Molecular phylogenetic studies indicate that Cedrus is the only problematic genus of the six genera included in Abietoideae (Abies-Cedrus-Keteleeria-Nothotsuga-Pseudolarix-Tsuga) [15].
2. Geographical Distribution of Conifers
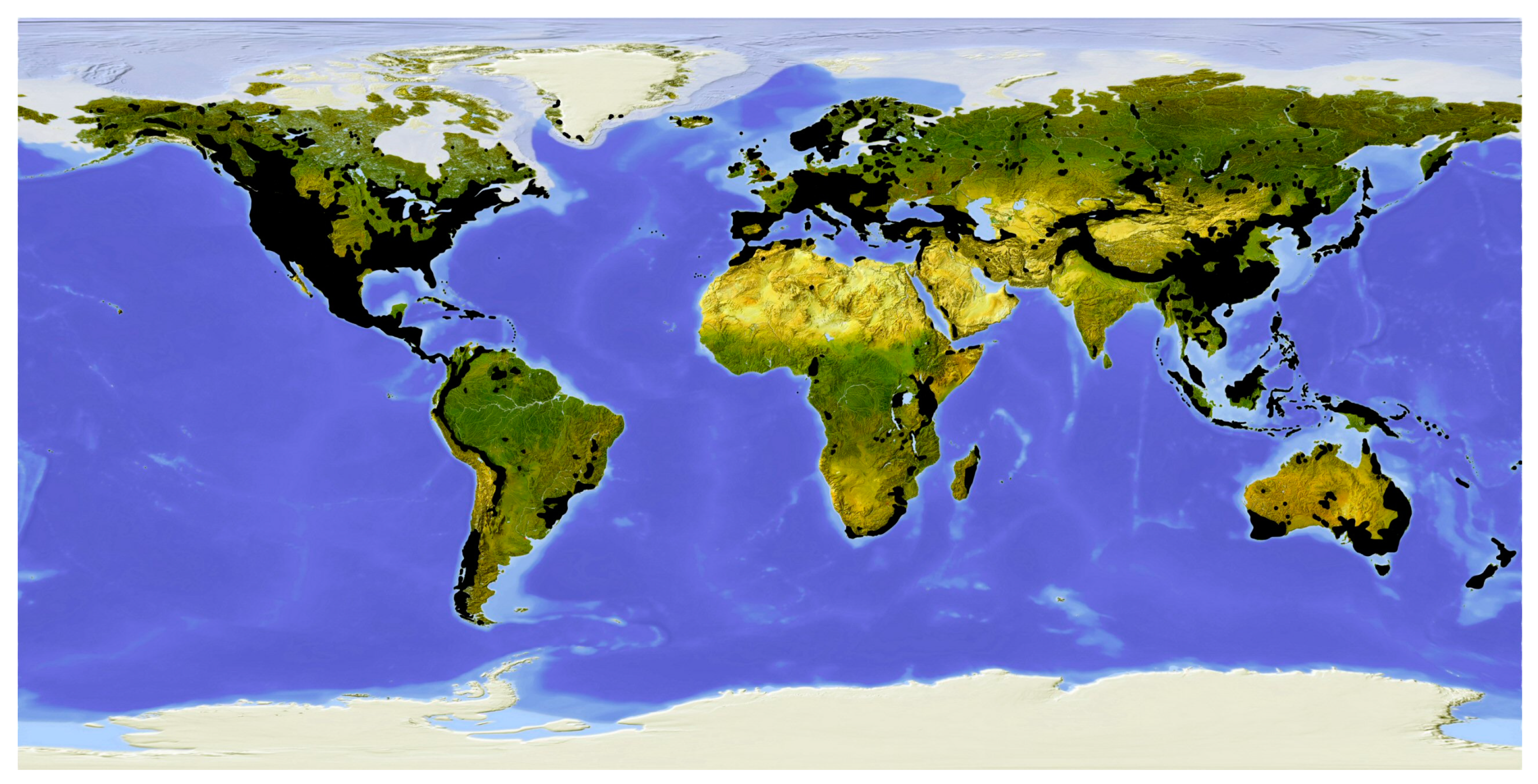
Conifers are widely distributed around the world and although they have been on Earth for more than 300 million years, they continue to occupy and dominate extensive areas of vegetation, particularly in the northern hemisphere.
The distribution pattern of conifers differs from north to south. The extensive boreal forest belt around Earth is dominated by a few species. The number of species increases in the south, with a predominant pattern of a more abundant presence of conifers in the principal mountain ranges, e.g., western North America, Mexico and Central America, the Andes, Europe (circum-Mediterranean forest belt from Spain to Greece), North Africa (Atlas Mountains), the Himalaya (extending towards China and Japan), New Guinea and New Zealand. In the eastern United States and southeast China, angiosperm forests predominate. Australia and Malaysia have unexpected occurrences of conifers, and some small islands are rich in conifers, e.g., New Caledonia, Taiwan and Tasmania. Conifers are absent only in Antarctica, the northern Arctic, steppes and deserts, the Amazon Basins, the Congo Basin and West Africa, where tropical forests occur, the Indian subcontinent (where only Nageia wallichiana occurs, in the Western Ghats), and most of the Tibetan Plateau. In Africa, in addition to forests in the north (Atlas Mountains from Morocco to Algeria), two other two geographical areas are significant for conifers: the region from Ethiopia to South Africa, and Madagascar [18] (Figure 6).
The families Pinaceae, Sciadopityaceae and Taxaceae are virtually confined to the northern hemisphere, whereas Araucariaceae and Podocarpaceae occur in the southern hemisphere and Cupressaceae occurs in both.
The extensive distribution of Podocarpus, Araucaria and Agathis in the southern hemisphere is thought to be due to the existence of the precursor species before the separation of the continents more than 125 million years ago [19].
3. Softwood Structure
3.1. Literature
Conifer wood anatomy has been extensively studied since the second half of the 19th century. Sanio [20,21] described compression wood for the first time, Castellarnau [22] studied the wood of Abies pinsapo, Kleeberg [23] studied conifer rays and Penhallow [24] proposed a classification of North American conifers. Further studies in the first half of the 20th century increased knowledge of softwood anatomy, including those by Jeffrey [25] on the comparative anatomy and phylogeny of Coniferales and extending knowledge of the genus Sequoia; Bitting [26] on histological differences between Pinus taeda and P. palustris; Bailey [27] on the wood structure of Pineae; Brooks [28] on the wood of Podocarpus spinulosus; Baker and Smith [29] on the pines of Australia; Thompson [30,31] on the origin of rays in conifers and ray tracheids in the genus Abies, respectively; Gordon [32] on the ray tracheids of Sequoia sempervirens; Holden [33] on ray tracheids in Coniferae; Chrysler [34] on rays in the genus Cedrus; Jeffrey [35] on the anatomy of woody plants; Kanehira [36] providing an anatomical description of Formosan woods; Kanehira [37] describing the anatomical features of the most important woods of Japan; Saint-Laurent [38] on the anatomical features of the woods of Algeria; Patton [39] on the anatomy of the Australian conifers; Pool [40] on the wood anatomy of Araucaria; Metcalfe [41] describing the anatomy of Fokienia hodginsii; Brown and Panshin [42] in their work “Commercial timbers of the United States: their structure, identification, properties, and uses”, extended in the 1940 edition; Brem [43] proposing a method to differentiate Picea and Larix; Bannan [44] on axial resin canals in Abietineae; Peirce [45,46,47,48] describing the xylem of Pseudolarix and Sciadopitys, the anatomical interrelations in Taxodiaceae, and the systematic anatomy of the wood of Cupressaceae, respectively; Shimakura [49] describing the wood anatomy of Taiwania; Bernath [50] on the conifers of Chile and Covas [51] on the native conifers of Argentina.
The work by Phillips [52], subsequently reprinted without changes (1948, 1959 and 1963), was the first publication about the wood anatomical features of conifers, providing detailed descriptions of their features, with instructions on how to interpret them where necessary, and constituting a seminal work for later studies [5,53,54,55,56,57]. Adopting the approach of the work by Clark [58] on hardwoods, it included a system of multiple-entry-perforated cards. This work can be considered a turning point both in describing conifer wood using terminology that is still used today, and in identifying conifer wood using a multiple-entry system. From its publication until the proposal by the IAWA Committee [5], the number of related works increased (e.g., [53,54,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88]).
3.2. Softwood Identification
The work by Phillips [52] “Identification of coniferous wood by their microscopic structure” is probably the turning point from which softwood features were grouped. Alongside the use of perforated cards, a qualitative leap occurred in softwood identification based on anatomy. The work described 33 features divided into five groups (general features, tracheids, parenchyma, rays and resin ducts), in a radical shift from traditional dichotomous keys to a multiple-entry key. It did not include biometry, except for the number of resin canal epithelial cells and cross-field pits. Greguss [53] conducted a broad descriptive study in his work “Identification of living Gymnosperms on the basis of xylotomy”, describing 345 species, and added 160 species in 1972 (Xylotomy of the living conifers). Subsequently, Esteban et al. [56] published “Anatomy and identification of conifers wood as a species”, proposing a multiple-entry identification key with 81 anatomical features (axial tracheids, 29; axial parenchyma, 8; rays, 37; resin canals, 7), including the geographical distribution and descriptions of 352 species. Rather than addressing the impossible task of identifying species, the authors aimed to characterise the species they listed. The features proposed were a little different from those addressed by the IAWA Committee [5] “IAWA list of microscopic features for softwood identification”, now the main document covering all softwood features available to be used by researchers to describe softwoods. Heinz [57] wrote the doctoral thesis “Systematische Erfassung und Dokumentation der mikroanatomischen Merkmale der Nadelhölzer aus der Klasse der Pinatae”, a broad study of softwood anatomy at family, genus and species level.
The use of computer databases, such as DELTA (Descriptive Language for Taxonomy), developed in Australia [90], and the addition of the conifer database to the University of North Carolina’s InsideWood platform in 2020 [89], have provided systems based on interactive multiple-entry keys that are very versatile for softwood identification.
Despite the anatomical homogeneity of softwoods, their structure contains sufficient features for reliable identification on a genus level. The objective of this study was to perform a literature review of softwood anatomy, which, alongside the contributions made by the authors, enables features with diagnostic value to be assigned to family and genus levels.
4. Material and Methods
The material comprised microscope slides of 352 species from the wood collection of the Universidad Politécnica de Madrid (UPMAw) and 420 species used by Heinz [57] in his doctoral thesis. All the samples were either collected by the authors themselves or obtained through exchanges with leading collections from highly recognised institutions dealing in wood anatomy, whose collections have been built up over decades.
Observations and photos were taken with a Leica DFC420 camera and Leica DM2500 light microscope (Leica Microsystems Inc., Buffalo Grove, IL, USA) and a JEOL JSM-6380 scanning electron microscopy (SEM).
For nomenclature and taxonomy at the family, genus and species levels, POWO [2] was used as the database. We also referred to The Gymnosperm Database. http://www.conifers.org (accessed on 20 November 2022) [91]. Only three exceptions were made: the genera Actinostrobus and Neocallitropsis were maintained as distinct from Callitris, and Cupressus and Hesperocyparis were dealt with together.
5. Microscopic Features
5.1. Axial Tracheids
One of the most unique features of conifer woods is the homogeneity of their microscopic structure, which is primarily composed of axial tracheids. Because of this, they are known as homoxylous woods.
Modern tracheids appeared in the Early Devonian, while ray tracheids and axial parenchyma have been present since at least the Late Devonian or Early Carboniferous [92].
Tracheids are the most abundant element in present-day conifer wood, comprising 90% of the total xylem volume [93]. The remaining 10% is made up of three elements: axial parenchyma, rays and resin canals. In Araucariaceae, the absence of axial parenchyma and resin canals results in a percentage of axial tracheids as high as 95%. Their typical shape is a long prism, with tips obliquely truncated by planes parallel to a line in the radial section. They are joined longitudinally through these planes, forming alignments parallel to the axis of the trunk [94].
With regard to tracheid length, in the Late Carboniferous some plants contained tracheids several centimetres long (Medullosa), although today they are barely more than 1 cm in length. This large size, which is associated with a high number of pits, has led some researchers [95] to suggest, in Carboniferous genera (Medullosa, Callistophyton and Lyginopteris), atypical hydraulic activity linked to warm humid environments with no risk of embolism caused by frost or drought. Strangely, although conditions were similar in some stages of the Mesozoic, no fossil records have been found with tracheids of that length or as many pits [96].
Tracheid length now varies from 2000 μm in Taxus baccata or 3500 μm in Larix decidua to 10,000 μm in Araucaria angustifolia. It is measured in macerated wood or using microscope slides using methods such as those described by Ladell [97] and Wilkins and Bamber [98], which are less accurate than maceration but provide satisfactory results. In any case, tracheid length must always be measured in mature wood due to the influence of the cambial age on length. The provenance of wood also affects tracheid length, with significant variation depending on whether the sample is from the trunk, branches or roots.
In some taxa with very low or very high tracheid length values, length can aid diagnosis, but in most cases it is of little diagnostic value.
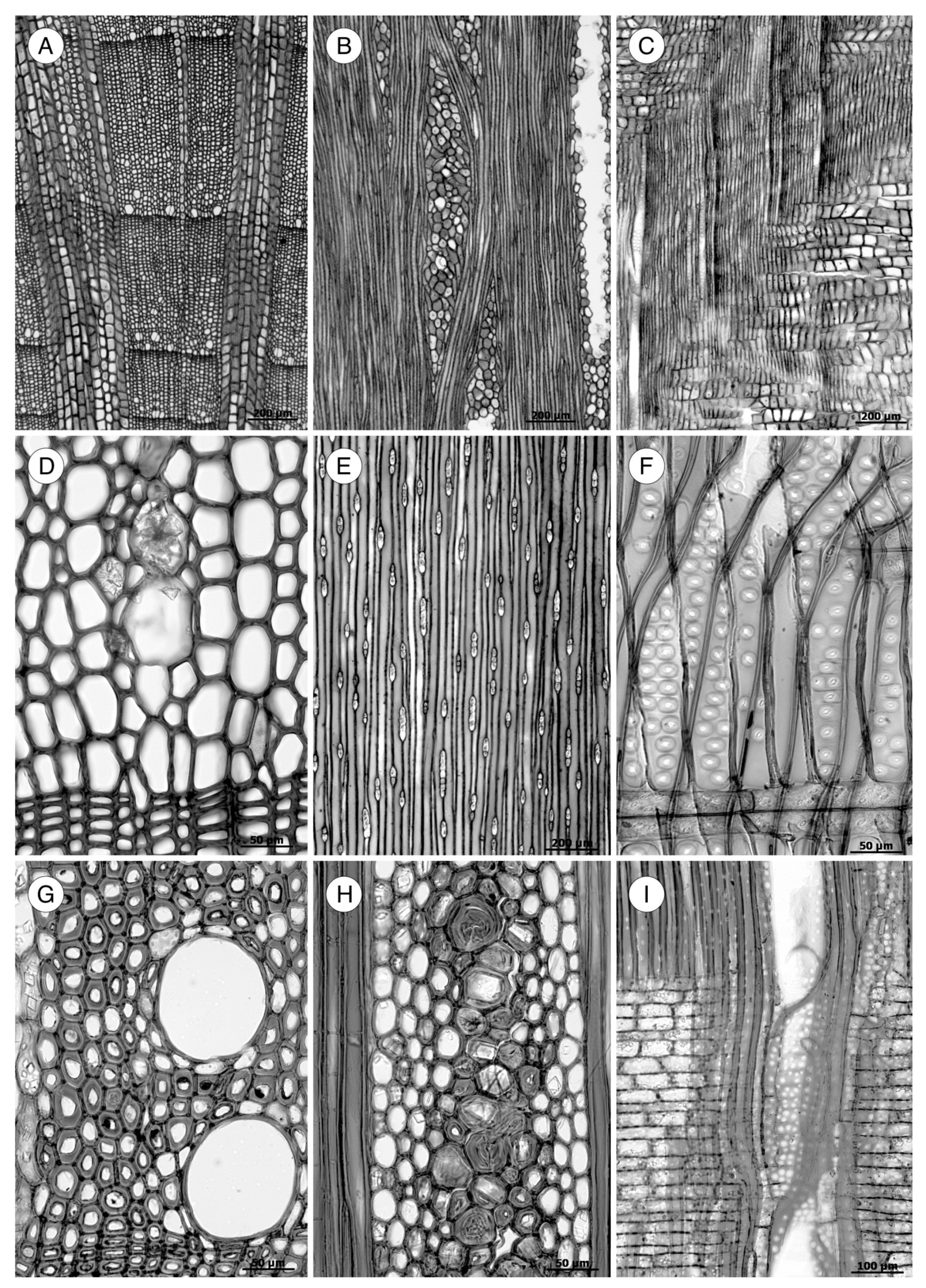
In the transverse section, trees from temperate forests in both hemispheres produce wood with true growth rings, showing differences between earlywood and latewood. Species of Cupressaceae, Pinaceae and Taxaceae in these areas typically produce wood with distinct growth ring boundaries. In tropical and subtropical forests, however, rings correspond to the rainy season and are indistinct or only slightly distinct. In tropical mountain sites, growth rings can be distinct or indistinct. Araucariaceae and Podocarpaceae species are characteristic of these areas. The transition from earlywood to latewood can be abrupt or gradual (Figure 7).
Rings are normally gently curved in their outline, but in some species they are undulating (Taxus spp. and Juniperus spp.) or very undulating, as in Pilgerodendron uviferum (Cupressaceae). Undulations must be differentiated from what is known as hazel growth, characteristic of some species such as Picea abies and Pinus jeffreyi, an unexplained phenomenon [57]. All these features are associated with differences between earlywood and latewood, growth ring type and transition must be interpreted with caution when identifying species because they are not constant features but depend on the location of the species. The same occurs with the presence of false growth rings, normally produced by a particular phenomenon, or wedging growth rings, which result from partially absent wood formation in the transverse section, due to, among other causes, insufficient supply of reserve metabolites [99,100] (Figure 8).
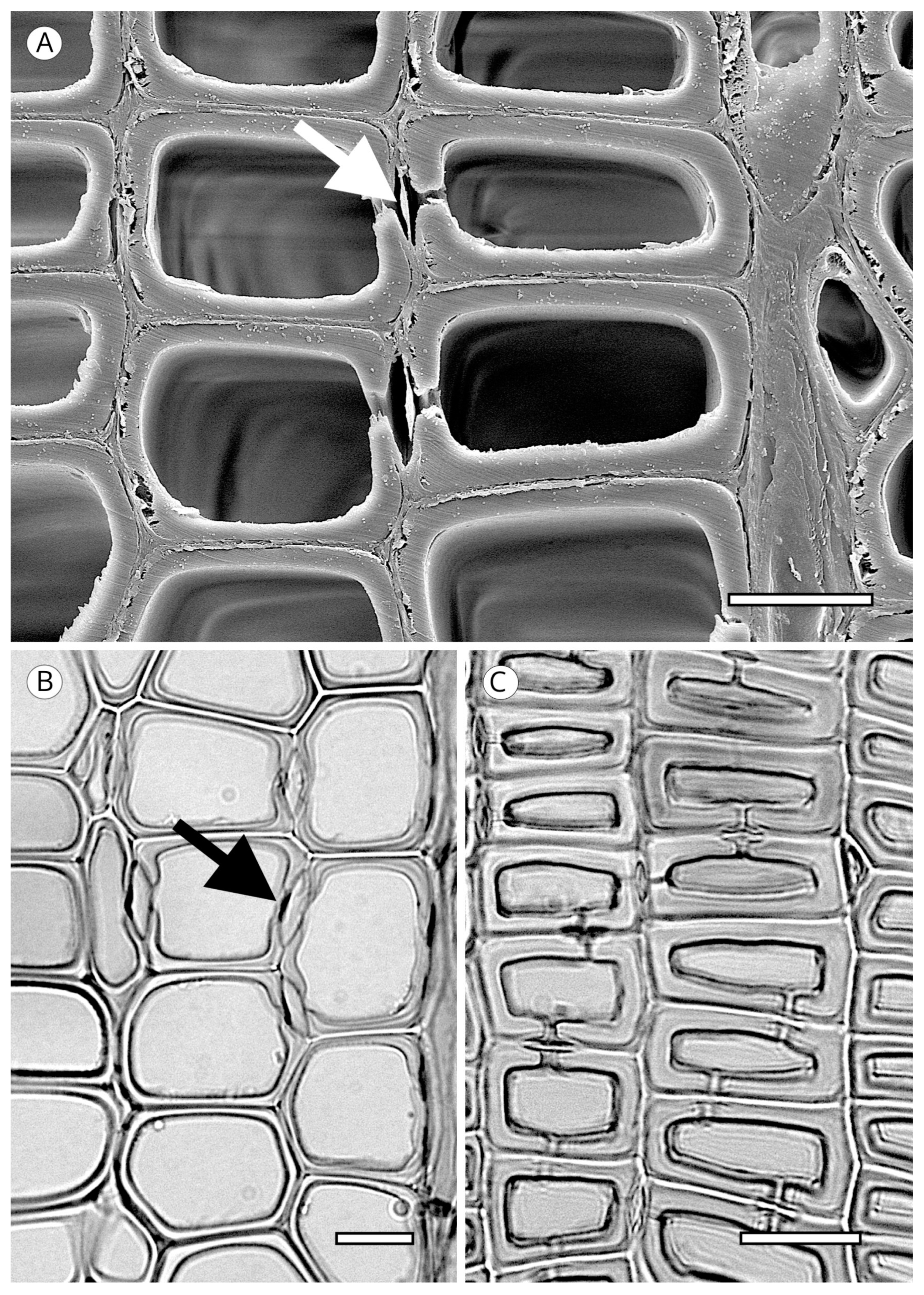
Although it is typically very laborious to obtain tracheid length, wall thickness is easily determined. The IAWA Committee [5] and Heinz [57] established two groups of wood based on the ratio of the double wall thickness (2WT), always measured in the radial direction, and the lumen diameter (L) in latewood tracheids: thin-walled (2WT < L) and thick-walled (2WT > L) (Figure 9). Despite the relatively low diagnostic value of wall thickness, some groups of wood have 2WT–L ratios that can be used for diagnosis, such as the soft pines (thin-walled) or southern yellow pines, Larix spp. and Pseudotsuga menziesii (thick-walled).
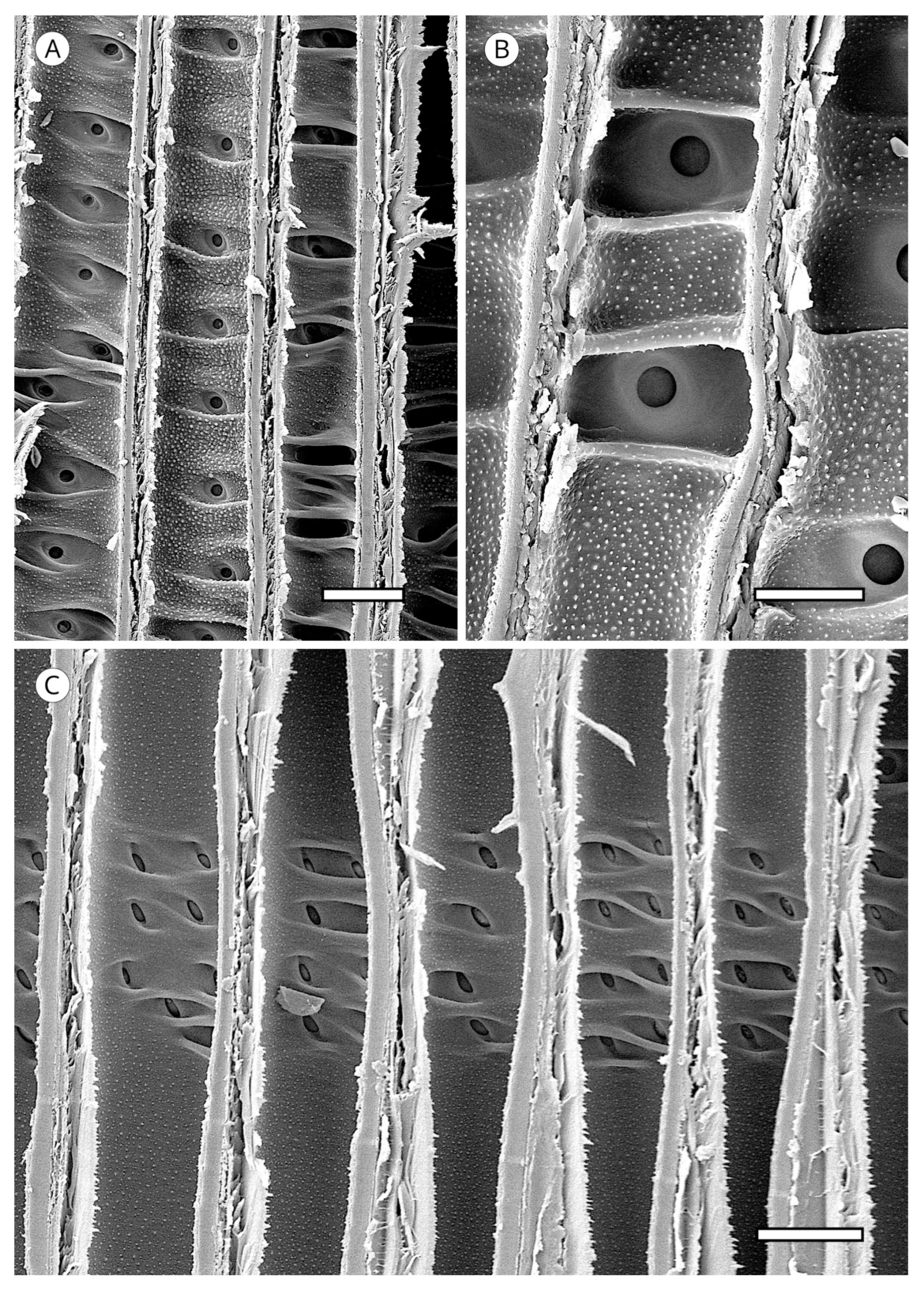
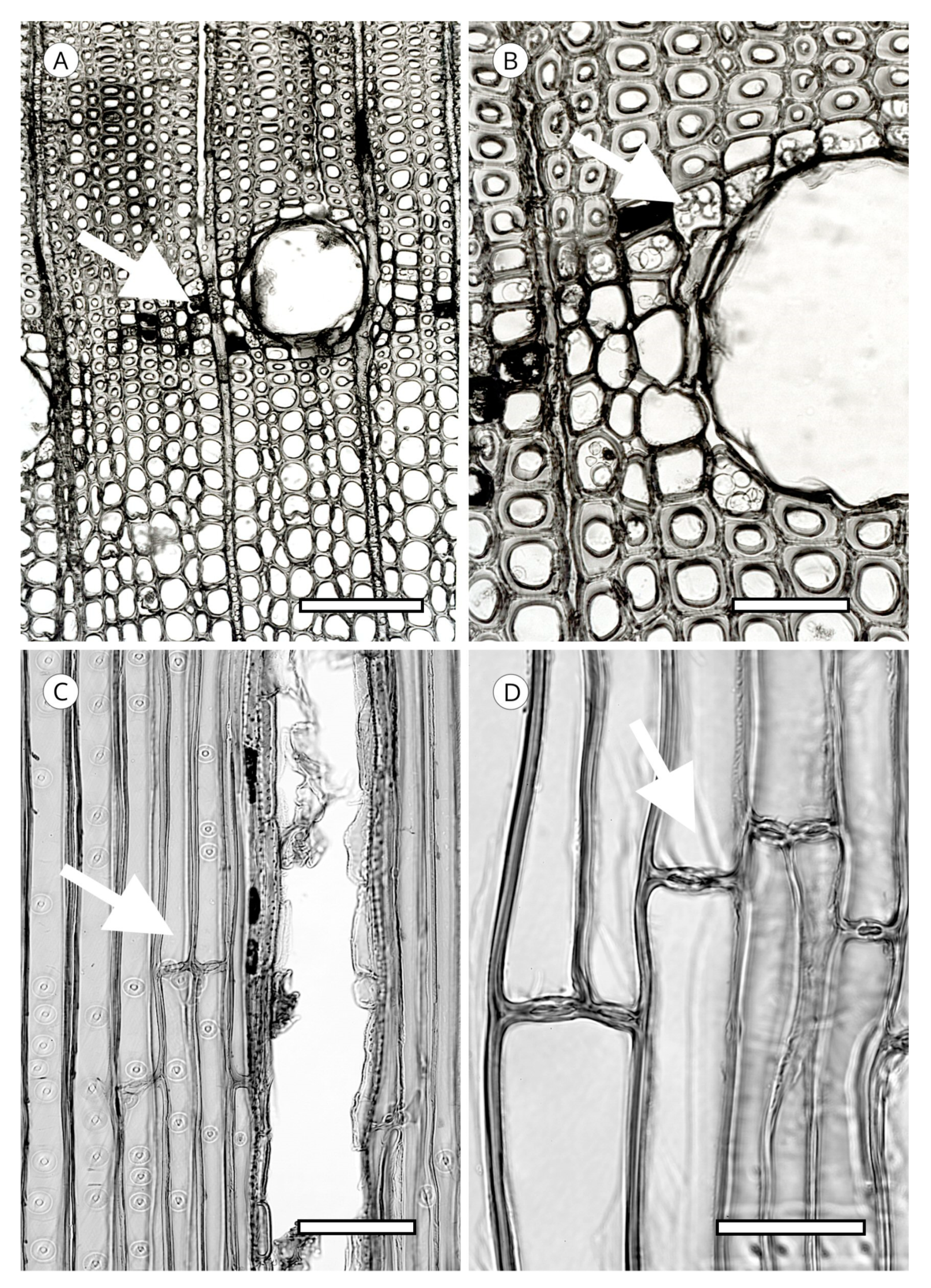
The transverse section of axial tracheids, associated exclusively with earlywood tracheids, is polygonal in most softwoods but circular in certain species, e.g., some species of the genus Juniperus. A third type described by Willebrand [101] in Agathis and Podocarpus, refers to axial tracheids with polygonal outline and circular lumen (Figure 9).
In some taxa, the transverse section may present intercellular spaces frequently associated with tracheids more or less circular in outline, although they are also associated with polygonal tracheids, e.g., Agathis alba and Pilgerodendron uviferum. This feature, characteristic of some taxa such as Juniperus communis, J. virginiana and Calocedrus formosana [4], must not be confused with the intercellular spaces generated in compression wood as a result of the circular tracheids generated in this type of wood. It can be easily differentiated by verifying, in any section, the spiral grooves inherent to compression wood (Figure 10). The absence of intercellular spaces cannot not be used for diagnosis.
These longitudinal intercellular spaces are connected only with radial intercellular spaces [102]. Intercellular spaces are generally thought to have three functions: for gas exchange, as a conduit for storage of water and nutrients, and as a pathway for diffusing heartwood substances [103].
5.1.1. Tracheid Pits
The first records of tracheid pits are from fossils of the Late Devonian–Early Carboniferous in progymnosperms and spermatophytes [104,105,106]. Although pits were originally circular, scalariform pits have also been found in tracheids of Rellimia and Protopitys [104].
The appearance of tracheid pits with a torus may have been one of the most important evolutionary stages in conifers. Their presence has ensured efficient conduction even in sites affected by drought and frost, safeguarding the existence of this type of plant in extreme climates such as tundra, deserts and high mountain sites, nearly always as dwarf trees and shrubs. Tracheid pits occur in all present-day conifers, in Gnetales and Ginkgoales but not in Cycadales [62], and have even been found in ferns of the genus Botrychium [107]. The earliest records are from the Middle Triassic [108] and their appearance remains an enigma in plant evolution, although they most likely correspond to the need to optimise conduction safety and efficiency.
Tracheid pits are located in tangential and radial walls, not always occurring in the former but always present in the latter, where they are larger in diameter and more numerous. Tangential wall pits are generally associated with latewood tracheids on the growth ring boundary and their presence appears to correspond to the improved transport at the end of the growing season (Figure 11). Unlike radial wall pits, they have no crassulae thickenings [93]. The greater spacing between tangential pits and their smaller diameter probably make strengthening the periphery of these pits with crassulae thickenings unnecessary.
Tangential pits have no diagnostic value. In a study of 230 species, Willebrand [101] concluded that the presence of this feature could not be assigned to any of them.
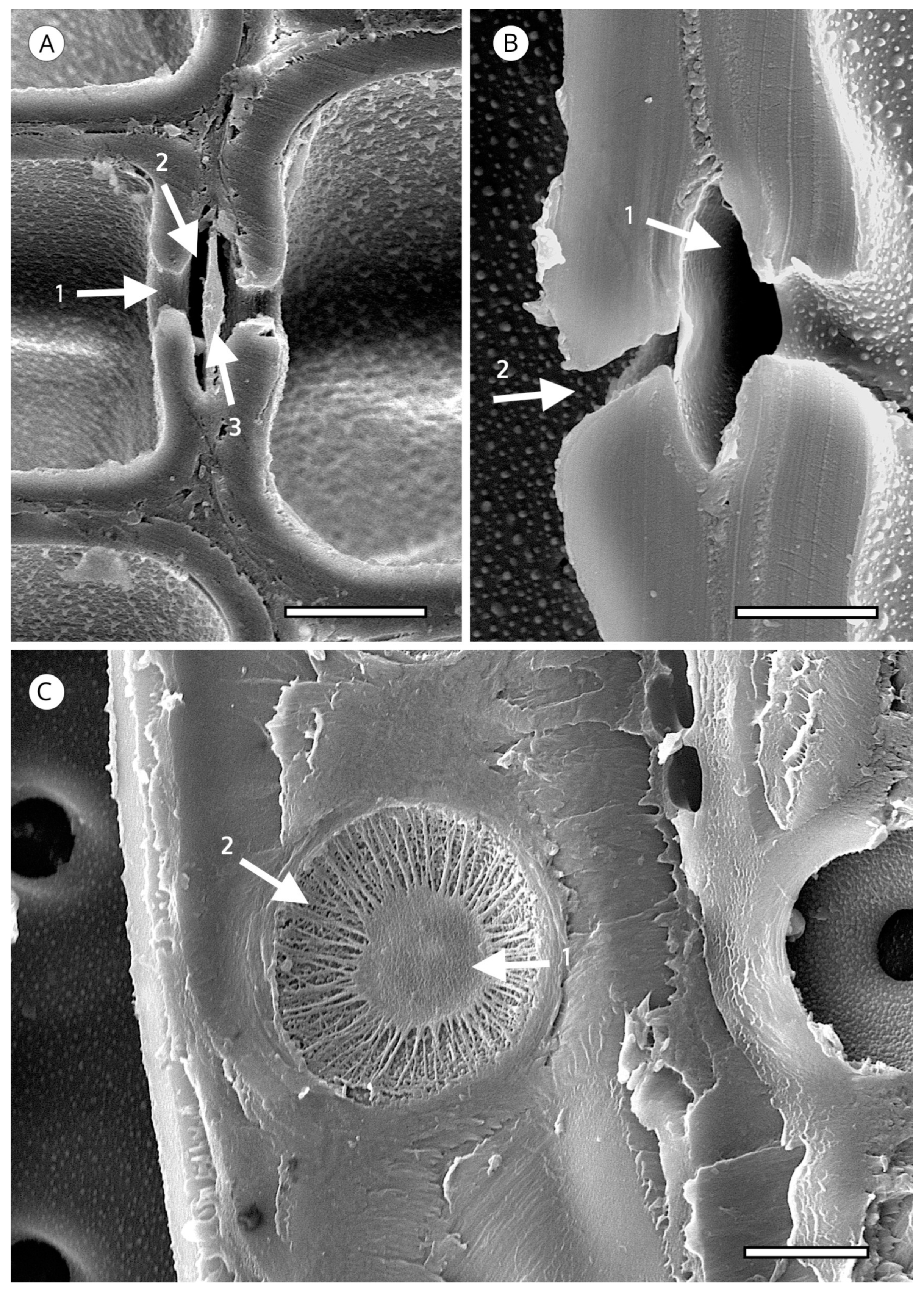
Tracheid pitting appears as a thickening of the secondary cell wall, extending over the non-thickened part in a dome shape, normally smooth and occasionally with radial striation, perforated at the vertex (aperture), leaving a space known as a chamber. The middle lamella has a thickening, known as a torus. Under normal conditions, water moves from one tracheid to another through the margo, which is located in the central plane of communication between the two tracheids and allows fluids to pass through it. When conditions are adverse, e.g., embolism due to extreme temperatures, the margo membrane is aspirated against the pit aperture and the torus plugs it, preventing air-seeding. Tracheids are not only efficient conductors, but also ensure a high level of hydraulic safety in adverse conditions. The torus has traditionally been thought of as an impermeable disc that functions as a valve, but in some species the torus is punctured by plasmodesmatal pores with a mean diameter of about 50 nanometres. Although most pores do not extend through the entire torus, there appears to be a relationship between species with a punctured torus and lower resistance to cavitation. However, it seems unlikely that punctured tori are the only cavitation mechanism in conifers, as this feature does not occur in all conifer species [109] (Figure 12).
The arrangement of tracheid pitting must be observed in earlywood tracheids. It is usually uniseriate but is biseriate in some taxa, generally opposite (e.g., Larix spp.). In some species of Pinaceae, tracheid pitting has a tendency toward alternate biseriate arrangement (e.g., Cedrus spp., Keteleeria spp.). Biseriate pitting is common in Araucariaceae, Cupressaceae (e.g., Widdringtonia, Taxodium), Pinaceae (e.g., Cedrus, Larix, Pinus, Tsuga) and Podocarpaceae (e.g., Phyllocladus, Podocarpus). Opposite multiseriate (three or more) tracheid pitting also occurs in Sequoia sempervirens, Taiwania cryptomerioides and Taxodium distichum [64]. In the three genera of Araucariaceae it is alternate and polygonal in appearance, as described by Phillips [52] in Araucaria and Agathis and Heady et al. [81] in Wollemia (Figure 13). Pit arrangement is a very important feature for wood identification and a key factor in plant evolution. Ancestral gymnosperm pits are thought to have been alternate [84].
The diameter of radial wall pits usually occupies two thirds of the axial tracheid diameter, although in Dacrydium and Podocarpus it can be less than half the tracheid diameter. This feature must be used with caution and only as a guideline, due to considerable intra-species variability [57] (Figure 13).
The IAWA Committee [5] divided the coniferous taxa into two large groups based on the torus in transverse section. In one group the torus is well defined, with a dense central area of the pit membrane with microfibril deposits in circular or radial orientation [110], often including incrustations with amorphous substances. In this group, the torus has a disc shape, visible in earlywood, a convex lens shape in latewood, or a flat transition from torus to margo. The other group comprises taxa without a defined torus, as in some species of Thuja and Thujopsis (Figure 14). The torus normally has a well-defined regular and smooth outline, but some taxa present variations that can be used to identify genus, e.g., Cedrus spp.
Cedrus spp. has scalloped tori (Figure 15), a feature characteristic of this genus. Despite reports of similar forms in Fitzroya [72] and transitional forms in Pseudolarix [101] and Cupressaceae [5], scalloped tori occur regularly only in all species of Cedrus, and therefore, this feature has diagnostic value. It is not known why Cedrus has this form of torus, but it could be associated with a strengthening of the margo for greater efficiency in situations of water stress. Liese [111] explained it as an accumulation of an irregularly dissolved matrix on the edge of the torus.
Cedrus has unique behaviour within Pinaceae, specifically in Abietoideae. According to Gernandt et al. [15], molecular evidence supports the genus Cedrus as a sister group to the other Pinaceae and should occupy a basal or root position in the family such that the subfamily Abietoideae can be considered paraphyletic. In addition to this special feature of Cedrus, the genus forms both axial and radial traumatic resin canals but lacks normal resin canals [14].
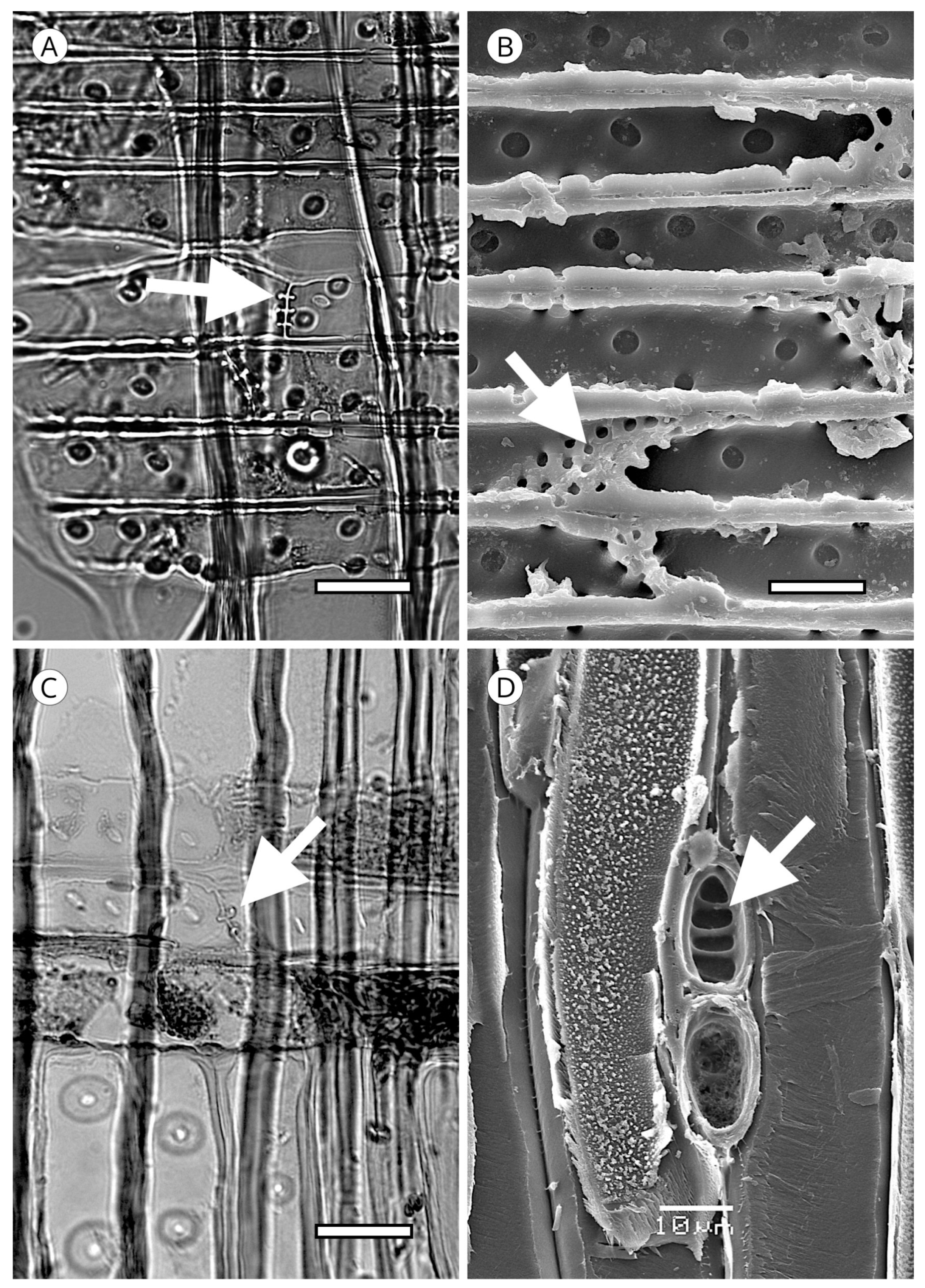
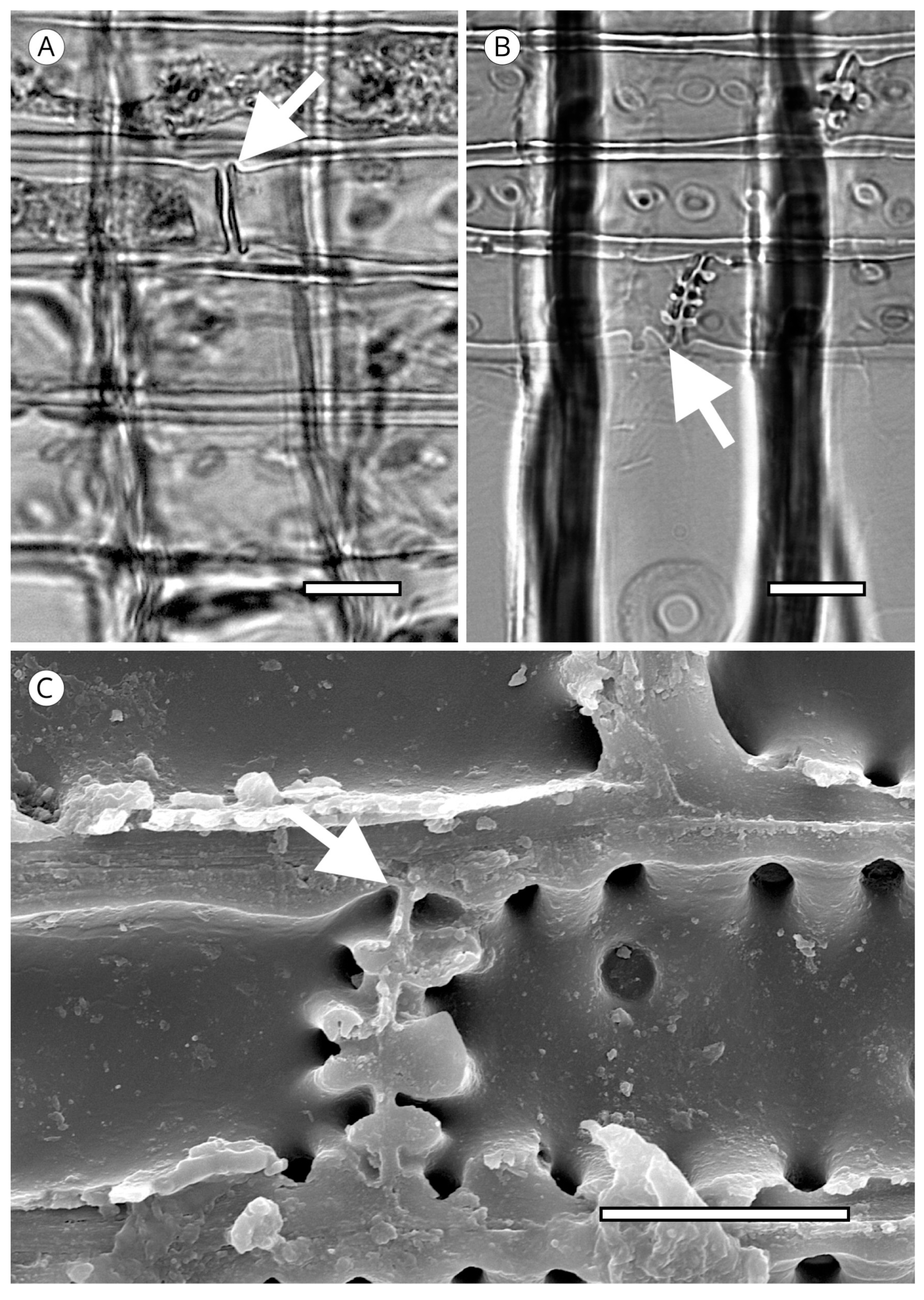
Another unique feature of the torus is torus extensions, or margo straps. These are radial thickenings from the torus to the margo periphery (Figure 15) made up of agglomerations of cellulose microfibrils. Like scalloped tori, they may correspond to the strengthening of the torus to improve the response to cavitation. Torus extensions occur regularly in Tsuga, Widdringtonia and Lagarostrobos franklinii [5], and more sporadically in Abies [112], Actinostrobus, Fitzroya, Juniperus, Pilgerodendron and Thujopsis [101]. In Abies, Heinz [57] confirmed the presence of extensions in 34 of the 48 species studied, while Esteban et al. [113] examined 33 species of Abies, one subspecies and four varieties, and confirmed the presence of extensions in 22 of them. Heinz [57] reported similar structures in Austrocedrus chilensis, Cupressus dupreziana and Juniperus spp., but the samples observed showed signs of degradation caused by fungi and/or bacteria, and therefore, this finding must be treated with caution.
In some species, the periphery of the pit has notched borders that can be used for diagnosis, particularly to separate almost identical taxa, such as Sequoia and Sequoiadendron, or even species of the same genus, as in the case of Juniperus [85]. They have been reported in Athrotaxis cupressoides, A. selaginoides, Chamaecyparis pisifera, Cryptomeria, Cupressus dupreziana, Juniperus thurifera, Papuacedrus papuana, Sequoia, Taiwania, Thuja occidentalis, Torreya californica, T. nucifera, T. taxifolia [101] and Juniperus brevifolia [85]. Heinz [57] recorded less pronounced notched borders in Athrotaxis spp., Juniperus foetidissima, Sequoiadendron, Thuja plicata, T. standishii and Thujopsis (Figure 16).
The presence of any of these three features associated with torus (scalloped tori, extensions and notched borders) therefore has significant diagnostic value.
5.1.2. Warty Layer
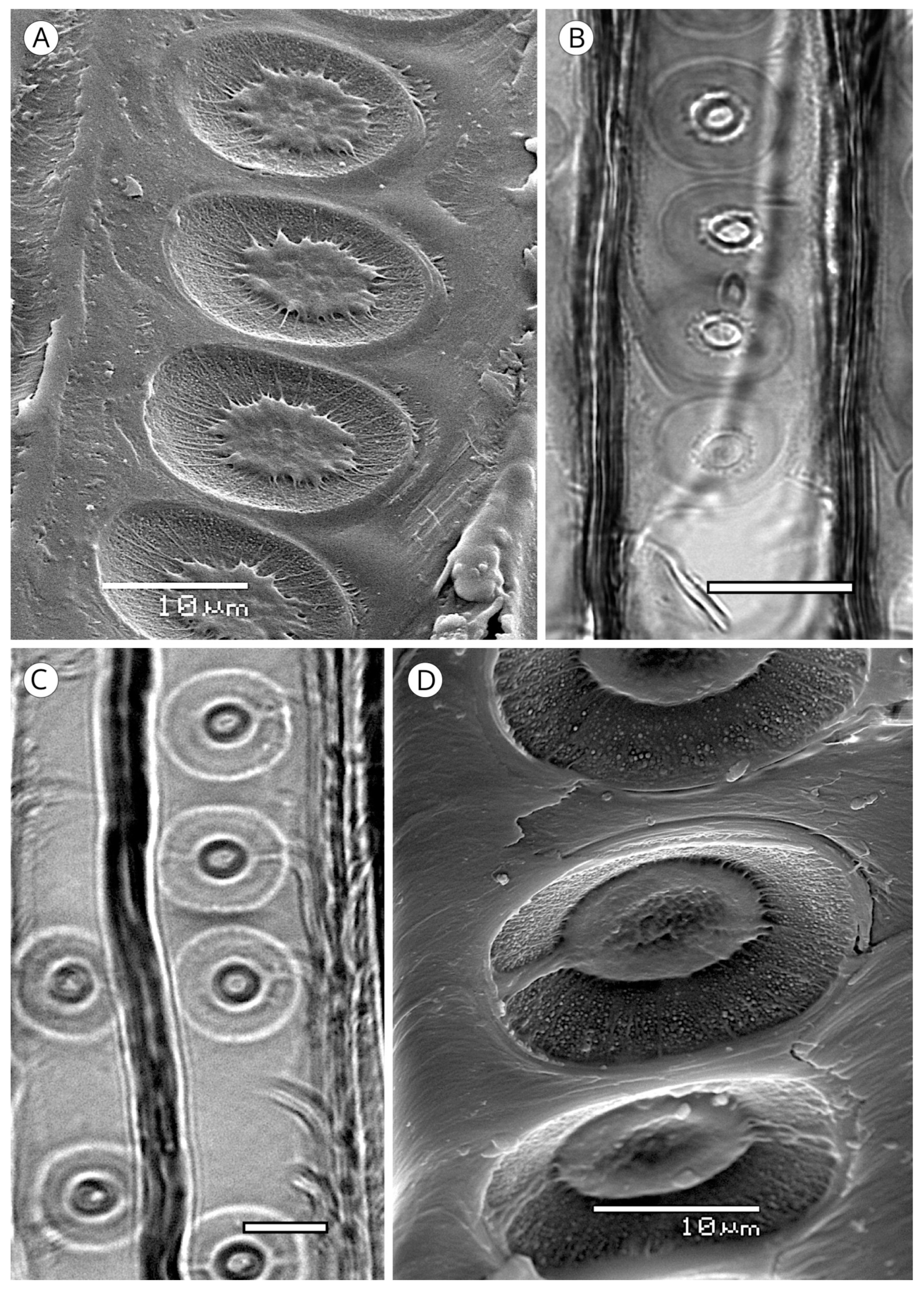
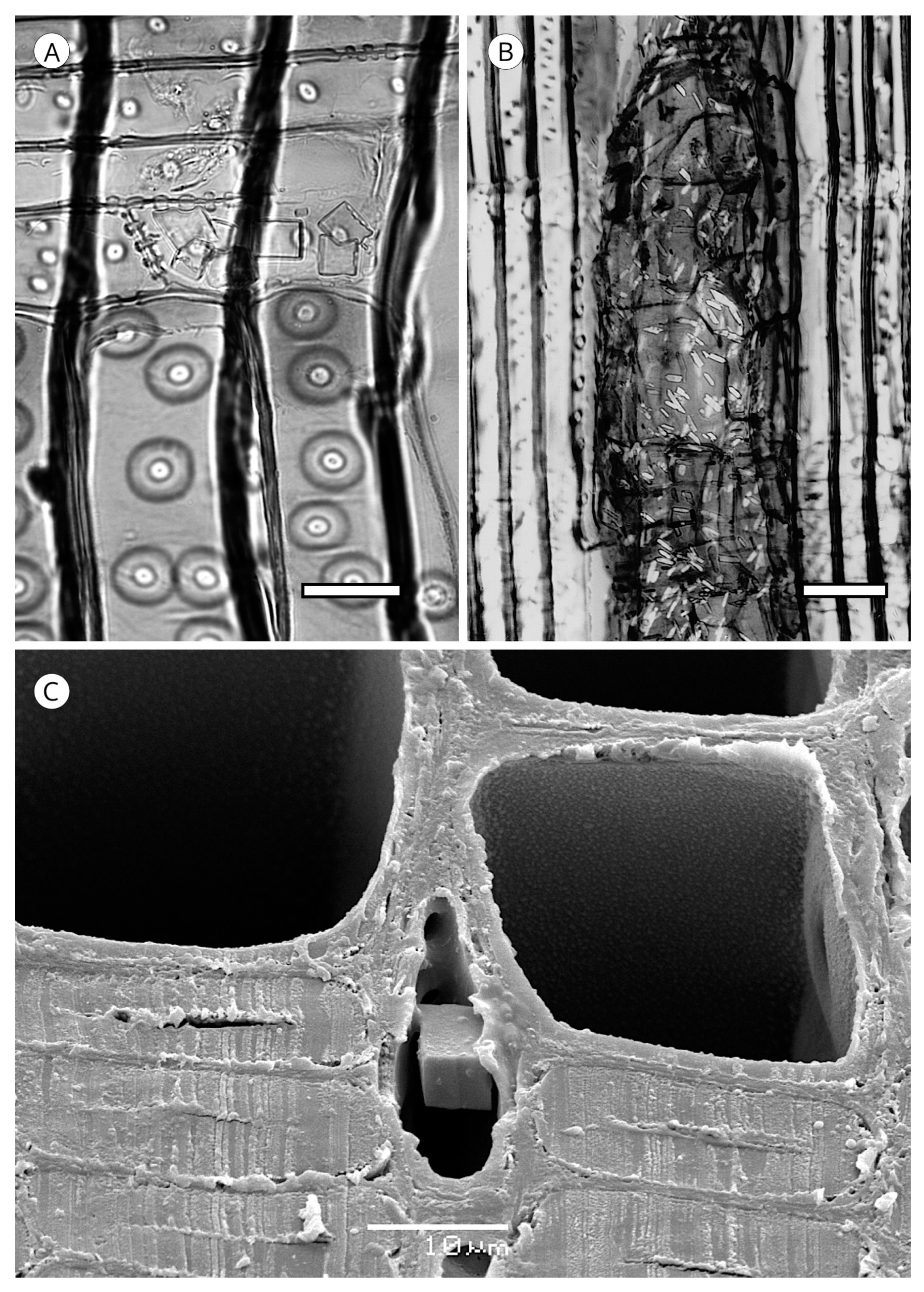
On the inner layer of tracheids (S3 or tertiary wall), some taxa present small wart-like thickenings 100 to 500 nm in diameter, rarely reaching 1 μm, and 500 nm to 1 μm in height, made up of lignin and hemicellulose and known as a warty layer. Liese [114] determined that the size, distribution and frequency of warts varies between taxa. Some authors have used this feature to compare genera and species, e.g., Heady and Evans [115] in Actinostrobus, and Esteban et al. [116] in Tetraclinis. The presence of a warty layer is a feature of significant diagnostic value.
Jansen et al. [117] reported that a warty layer is present in most conifer wood, but Harada et al. [110] noted its absence in Taxus cuspidata, Taxus floridana, Torreya nucifera (Taxaceae), Cephalotaxus harringtonia var. harringtonia, Nageia nagi and Podocarpus macrophyllus (Podocarpaceae). However, as Heinz [57] indicated, these observations are based on electron microscopy and few species have warts that are large enough or grouped densely enough to allow observation with light microscopy.
In some genera (Abies, Actinostrobus, Juniperus and Callitris) the warty layer can be observed without difficulty under light microscopy (Figure 17), but scanning electron microscopy (SEM) is recommended for biometry and morphology (Figure 18). Warts have been reported in numerous genera of Cupressaceae (Athrotaxis, Chamaecyparis, Cryptomeria, Cupressus, Fitzroya, Sequoia, Sequoiadendron, Tetraclinis, Thuja, Thujopsis and Widdringtonia), in Pinus (P. bungeana, P. massoniana) and some Podocarpus [5]. Heinz [57] reported them in Diselma, Fokienia and Platycladus (Cupressaceae) and in Pinus gerardiana, P. monophylla and P. quadrifolia (Pinaceae), and Esteban and de Palacios [14] in the subfamily Abietoideae (Pinaceae) (Abies, Cedrus, Tsuga).
Based on the hypothesis of Zimmermann [118], Heady and Evans [115] proposed that the small warts of slightly more than 1 μm in height in Actinostrobus are responsible for catching the air bubbles produced in recently thawed tracheid water to prevent them from coalescing and forming large bubbles that could create a risk of blockage in tracheids. Such bubbles have the potential to block the tracheids by expanding as xylem pressure decreases and transpiration restarts [115]. The three species of Actinostrobus have thin barks that are probably insufficient to counter the effect of extreme temperatures (especially frost) where they grow, and they frequently develop frost rings in their wood. Frost rings have been observed in other Cupressaceae, e.g., Diselma, which, strangely enough, has somewhat smaller warts. Other genera of Cupressaceae, characteristic of drier sites, such as Tetraclinis, also contain the same type of warts, probably to prevent cavitation by water stress, but the question remains as to why other conifers in the same or similar sites have no warty layer in their tracheids. The warty layer could, therefore, be a defence mechanism against cavitation in species subjected to extreme temperatures, but this can be confirmed only by physiological studies.
5.1.3. Organic Deposits
Some species develop organic deposits in heartwood tracheids (Figure 19). They are mentioned in the scientific literature as resin plugs, resin spools or resin plates, and although they are visible in the transverse section, they are more readily observed in longitudinal sections. They are normally located near the rays and are dark-coloured, occupying all or part of the cell lumen. Because of their unique features, their presence is of significant diagnostic value. Without naming them as such, Peraza [119] described organic deposits in the resinous wood of Pinus canariensis. They have been reported in axial tracheids in Abies pinsapo [120] and Halocarpus biformis [57], and described in Agathis and Araucaria, normally more abundant in the former.
Deposits in parenchyma cells have been described in Callitris glauca, Calocedrus spp., Cupressus spp., Fitzroya cupressoides, Juniperus procera, J. virginiana, Thuja plicata, T. standishii, Widdringtonia spp., Dacrydium elatum, Podocarpus totara, P. ferrugineus and Saxegothaea conspicua [55].
The presence of organic deposits in tracheids seems to be associated with the mechanisms of heartwood formation as a tree defence against attacks by xylophagous agents. When a tracheid cell lumen is plugged, hyphae progression is prevented. The occurrence of organic deposits in tracheids in wood without normal resin canals, e.g., Abies, Agathis and Araucaria, indicates that tracheid plugging cannot be explained solely by the presence of resin canals. The blockage of axial tracheids can be explained only by a metabolic pathway, such as that described by Hillis [121] for impregnating axial tracheids with resin in Pinus radiata through cross-field pits from the ray parenchyma cells.
It should be noted that when preparing microscope slides, particularly during at the dehydration stage, organic solvents may dissolve the content of deposits and cause the loss of this diagnostic feature. Additionally, tracheids in archaeological wood can resemble organic deposits due to encrusted extraneous substances such as paint, resin, tar, oil or iron oxide [57].
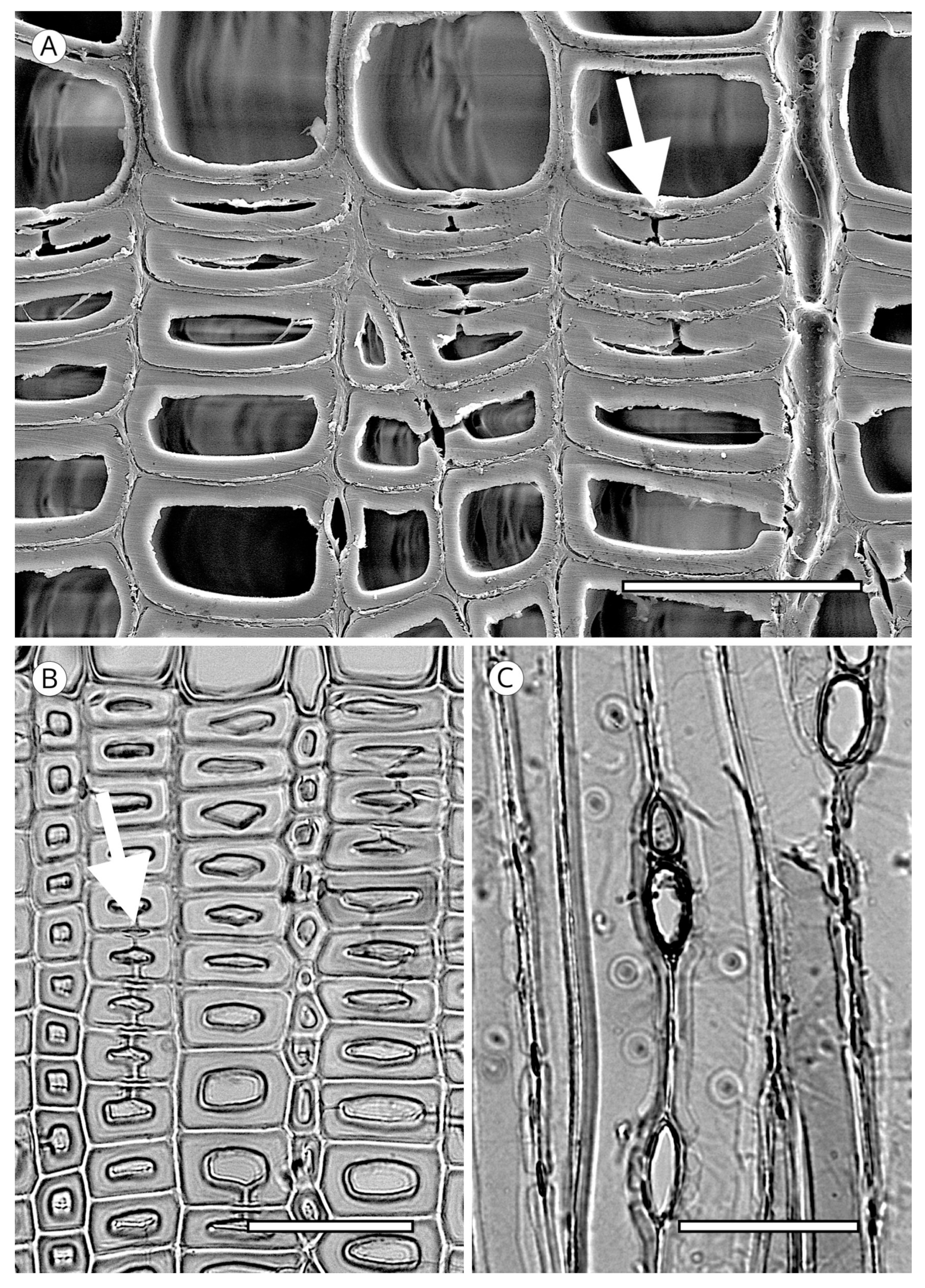
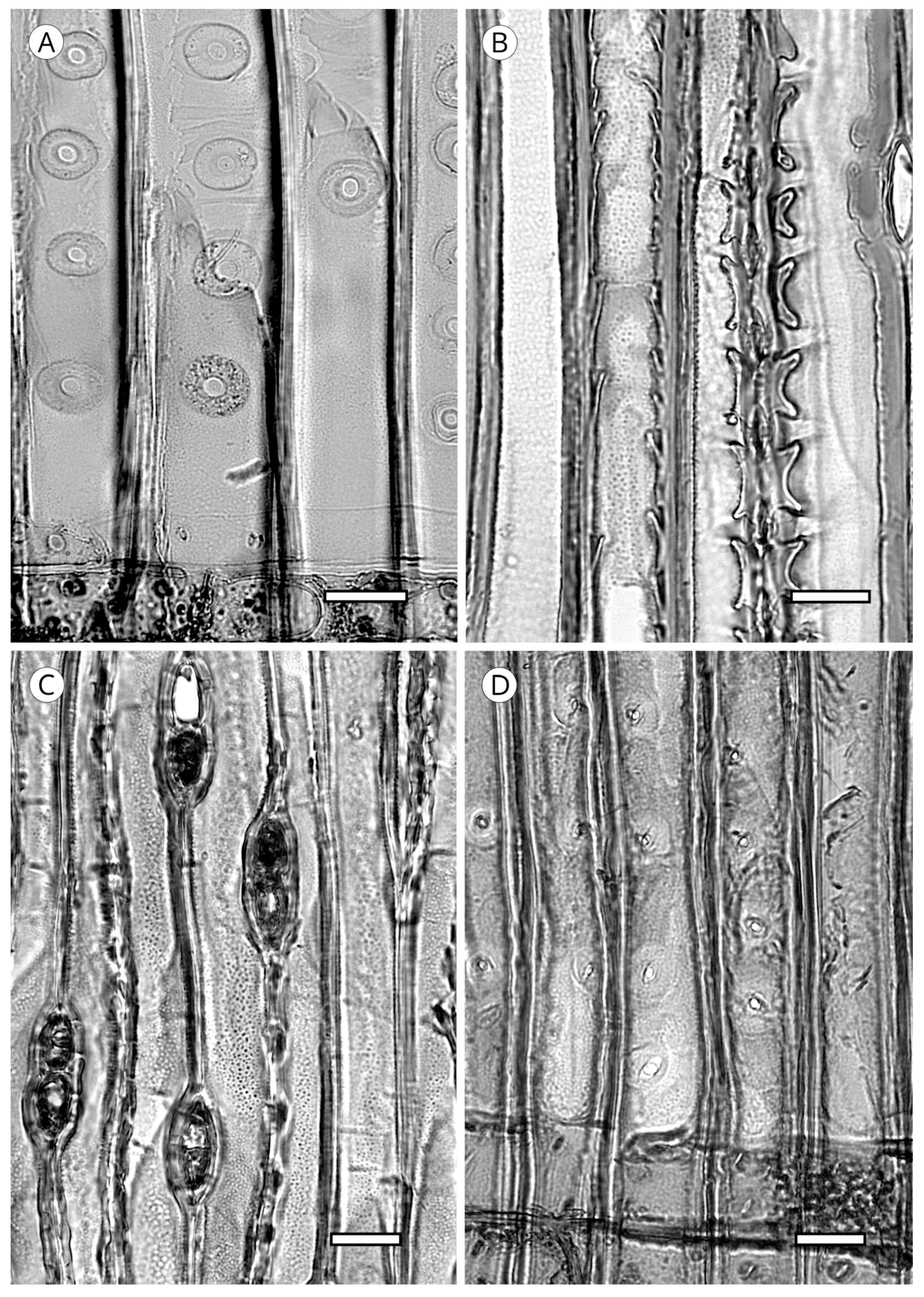
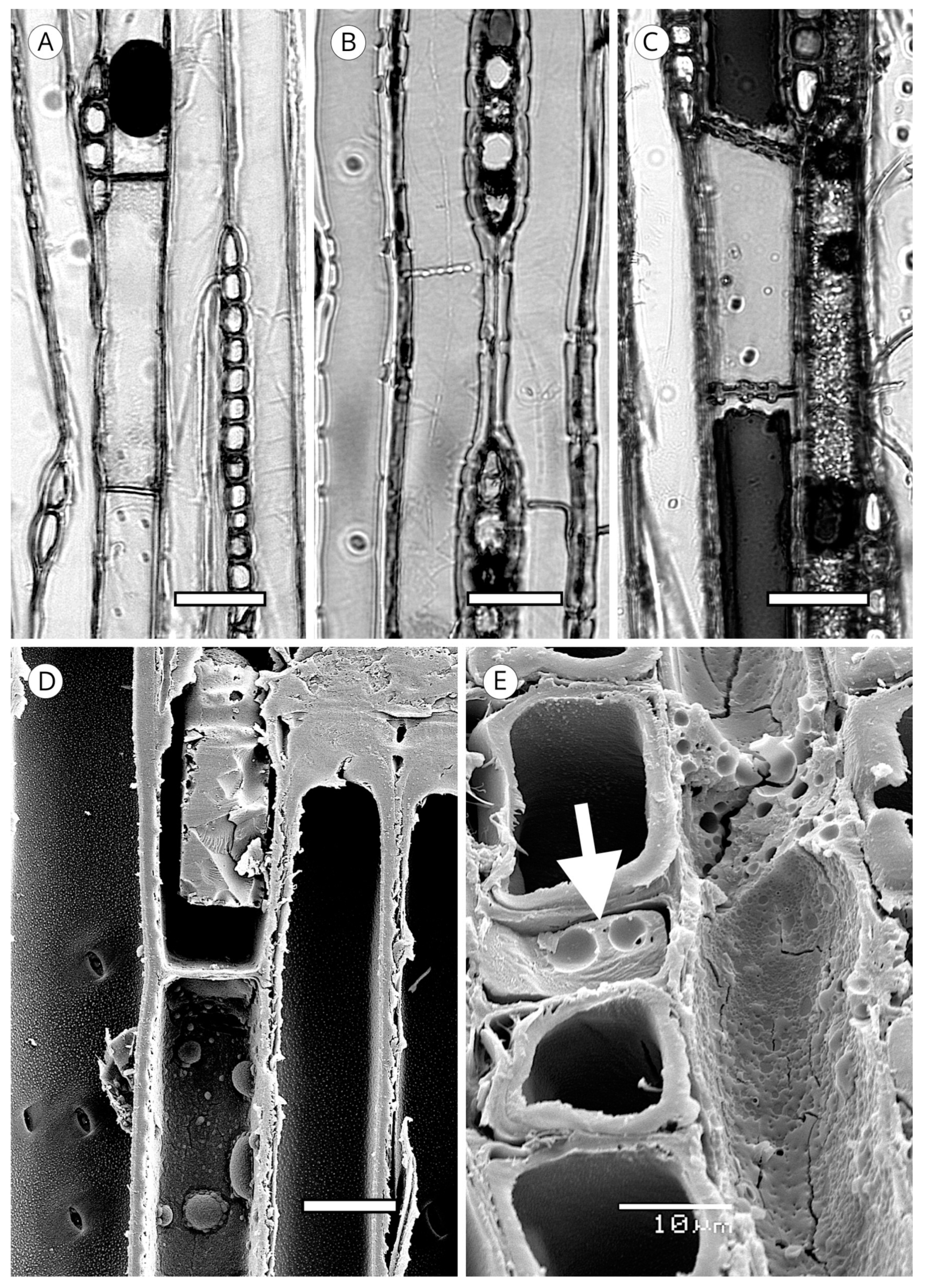
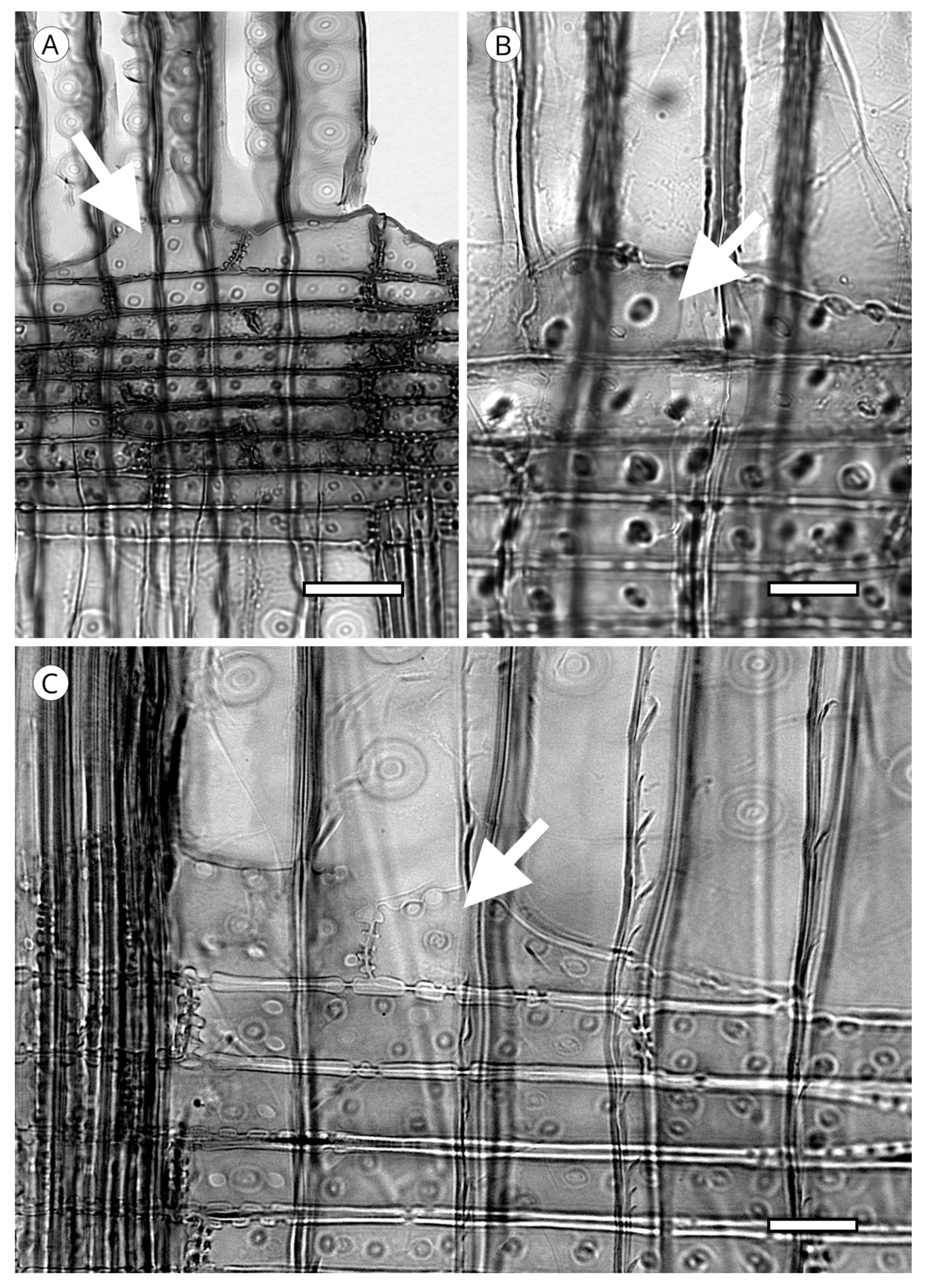
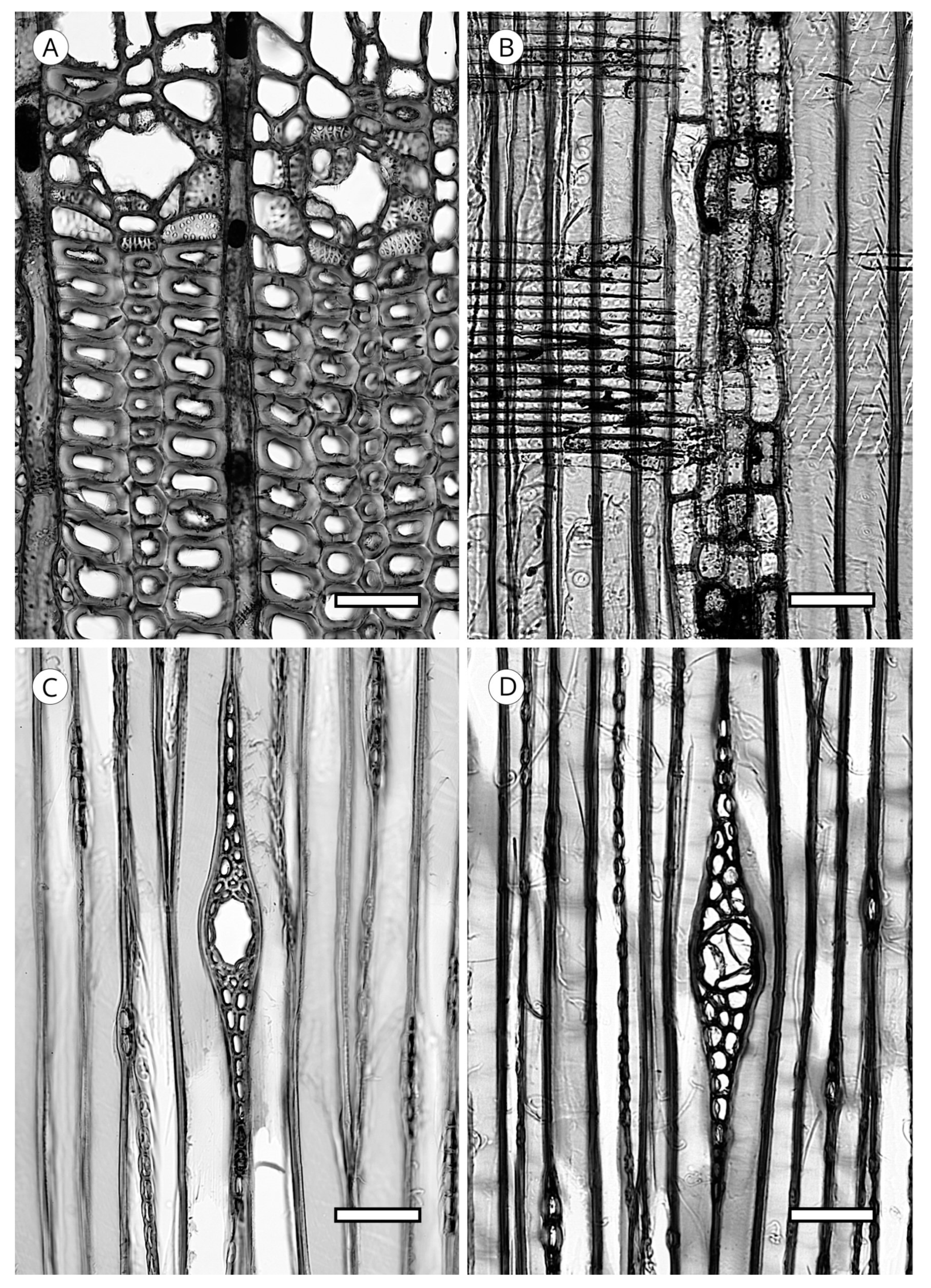
5.1.4. Helical Thickenings
Some species always have thickenings in the form of ridges that occur as helicoids on the inner face of tracheids (Figure 20). Helical thickenings are true deposits inside tracheids.
These ridges are produced during the final stage of secondary wall formation through cortical microtubules obliquely oriented to the axis of the cell [122].
They have been reported in all genera of Taxaceae (Amentotaxus, Pseudotaxus, Taxus, Torreya and Cephalotaxus) except in Austrotaxus [56]; in Cathaya, in earlywood and latewood [123,124,125,126,127,128]; in Pseudotsuga [56,125], in Pseudotsuga sinensis and P. menziesii in earlywood and latewood, although in the latter they are sometimes present only in latewood; in some species of Larix, only in latewood tracheids (L. decidua, L. griffithii, L. occidentalis, L. potaninii); in Picea spinulosa, well-developed in earlywood and latewood, and in Picea abies, P. brachytyla, P. glauca, P. glehnii, P. jezoensis, P. koraiensis, P. mariana, P. sitchensis and P. torano, only in latewood [129]. For other authors [60,130] they are regularly present in earlywood and latewood in Picea brachytyla, P. maximowiczii, P. morrisonicola, P. purpurea, P. schrenkiana, P. smithiana and P. spinulosa, later confirmed by Heinz [57]. Helical thickenings are most easily seen in earlywood because in wood with particularly narrow rings, they are either overlooked or simply not visible.
The grouping of helical thickenings (single, double or triple), spacing, inclination angle, thickness, branching and connection to the inner cell wall vary among taxa. Some parameters can be quantified, but others are highly variable and difficult to measure. In Taxus and Pseudotsuga, helical thickenings are normally single (Figure 20), whereas in Amentotaxus and Torreya they are double (Figure 20) or even triple in Torreya. In Cephalotaxus they can be grouped or not grouped in the same wood [57]. The inclination angle and thickness of spirals are correlated with spacing. Pseudotsuga and some taxa of Picea have very thin spirals with little spacing, forming an angle of 80° to 90° with the tracheid axis, while in Amentotaxus, Cephalotaxus, Taxus and Torreya they are typically thicker and have a steeper angle. The spacing between helical thickenings is considered narrow when there are typically more than 120 spirals per axial mm, e.g., Pseudotsuga (120–180) and Picea smithiana (150–200), and wide when there are typically less than 120 spirals per axial mm, e.g., in Torreya, with 80–100 (40–50 pairs), and Taxus (40–80). Cephalotaxus has 80–140, and may be included in either category depending on the species and/or sample [5]. Some authors [78] observed differences in taxa with helical thickenings in the connection of spirals to the S3 layer. For example, in Taxus, Torreya and Cephalotaxus, helical thickenings have a narrow base loosely connected to the inner layer of the secondary wall (S3) (Figure 21B), while in Pseudotsuga, they have a wider base, as if the spirals were more securely fastened to the S3 layer by connecting microfibrils (Figure 21A). Moreover, in some species of Picea, Torreya and Cephalotaxus, helical thickenings can be present in branches but absent in the trunk [131].
Spiral grooves found in compression wood must not be confused with helical thickenings. Spiral grooves are normally at steep angles of around 45° and appear as cracks in the cell wall (Figure 21C,D). The same can occur in archaeological wood, where the enzymatic activity of soft-rot fungi produces cavities resembling spiral grooves in compression wood [132] (Figure 21C,D).
The mechanical effects of helical thickenings on the cell wall are unknown. It appears that these ridges, with a small diameter relative to cell wall thickness, have little influence on the mechanical properties of the wood that contains them, although they may affect water conduction in the cell lumen [133].
The presence of helical thickenings and their grouping (single or in pairs or triplets) have significant diagnostic value.
5.1.5. Callitroid Thickenings
Another type of thickening present in the secondary wall of longitudinal tracheids is the callitroid thickening. It occurs in all species of Callitris and in the latewood of the three species of Actinostrobus [115], although it has also been reported in other taxa, e.g., Phillips [132] noted its presence in Pseudolarix and Juniperus, Howard and Manwiller [134] in some species of the southern pine group, Meylan and Butterfield [135] in Dacrydium cupressinum and Schweingruber [71] in Tetraclinis. However, according to Phillips [52], these observations should be considered “rare and inconspicuous”. This type of thickening must, therefore, be reserved as a feature exclusive to Actinostrobus and Callitris, and thus, its presence has significant diagnostic value.
Callitroid thickening appears as two parallel ridges above and below the pit aperture, resembling an awning in the tangential section. Heady and Evans [80] also recorded the presence of this thickening in cross-field pits (Figure 22 and Figure 23).
Heady and Evans [80] showed that the frequency of thickening is higher (>85%) in species of Callitris from dry sites in Australia (C. canescens, C. endlicheri, C. glaucophylla, C. tuberculata and C. verrucosa) than in humid sites with high precipitation (<10%) (C. macleayana, C. neocaledonica, C. oblonga and C. sulcata), and that in species with higher frequency of callitroid thickening, the thickening occurs in tracheids with a smaller diameter. Because of this, these authors suggested that the transverse bars on each side of the pit aperture, the region of the dome where the cell wall is weakest, could be regarded as reinforcement or supporting braces for the pit to prevent collapse.
5.1.6. Bars of Sanio
Bars of Sanio, or crassulae thickenings, are present in all conifer wood except Araucariaceae, where their absence was confirmed by Brown et al. [93] in Araucaria and Agathis and Heady et al. [81] in Wollemia. They have no diagnostic value. They are formed by a thickening of the middle lamella and the primary wall on the outside of both uniseriate and multiseriate pits and their mission seems to be to strengthen the cell wall on the periphery of the pits (Figure 24) [93]. This feature is not exclusive to conifers, as it has also been reported in Centaurodendron dracaenoides [136].
5.1.7. Trabeculae
Trabeculae occur occasionally in the axial tracheids of some conifers. They can occur in all conifer wood and have also been observed in hardwoods. They have no diagnostic value. They are cylindrical bars that extend through the tracheid lumen from wall to wall (Figure 25). Bars are usually wider where they join with the wall, and in species that have a warty layer, the warts extend through the bar of the trabecula. In earlywood trabeculae, they are usually thin, but in latewood they are shorter and wider. When trabeculae are present they do not usually appear in only one tracheid, but in several adjacent tracheids from the same cambial cell and at the same height. Because of this arrangement, [93] considered that trabeculae developed from the cambial cell. McElhanney et al. [137] attributed the formation of trabeculae to an accumulation of cell wall material on the hyphae of cambial fungi. Troncoso and Greslebin [87] demonstrated that trabeculae the formation in Austrocedrus chilensis wood is clearly associated with invasion by Phytophthora austrocedri.
Although Grosser [138] suggested that trabeculae act as bracing elements in tracheids to prevent collapse, this is unlikely in view of the scarcity and irregularity of their presence when they occur.
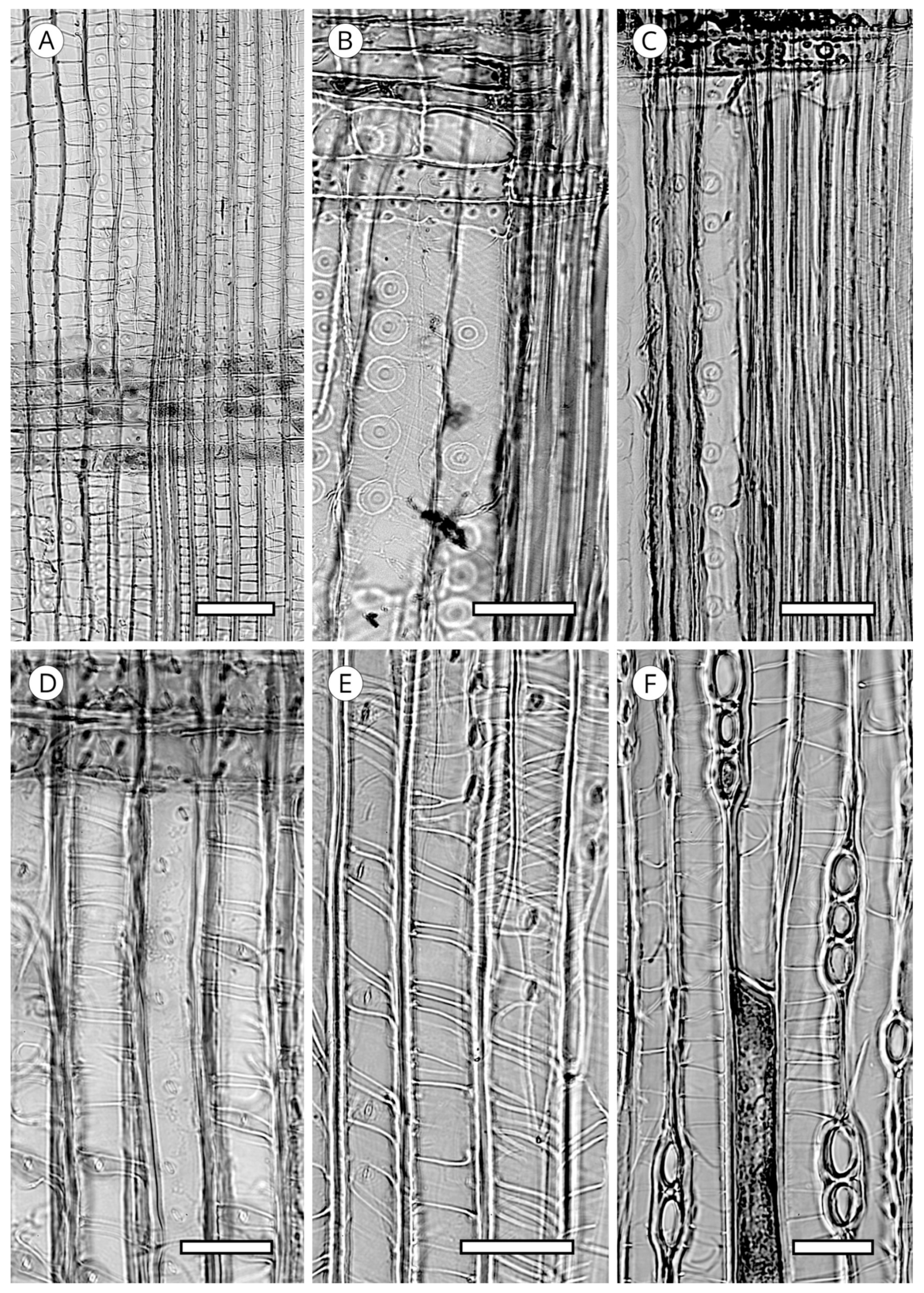
5.2. Axial Parenchyma
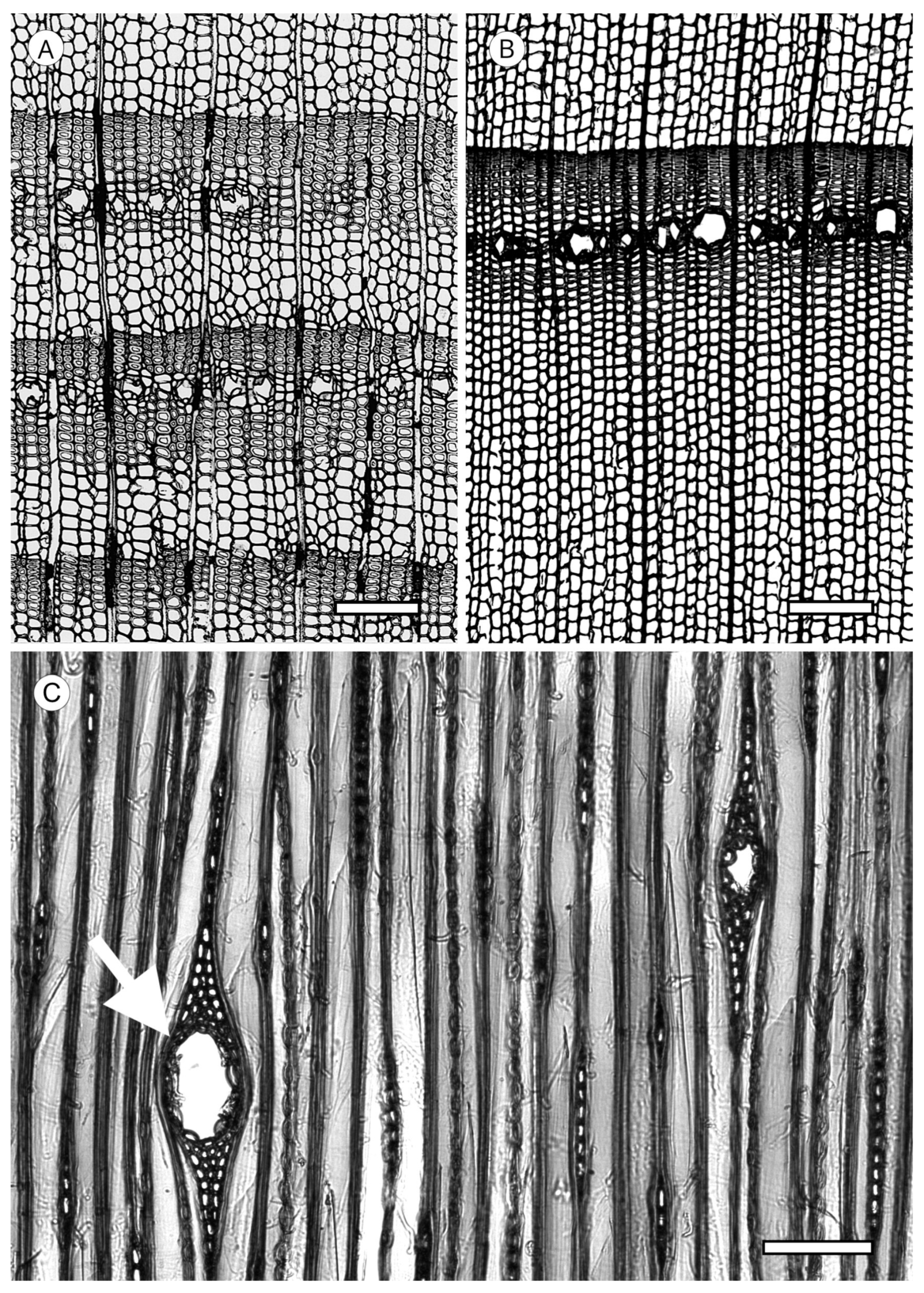
Axial parenchyma is less abundant in softwoods than in hardwoods. It appears in strands of more or less rectangular cells, generally with simple pits, that are responsible for storing metabolites. Its presence must be confirmed in longitudinal sections. It is rare or absent in Araucariaceae, Phyllocladaceae and Sciadopityaceae. Ghimire et al. [139] recorded it in Amentotaxus and Austrotaxus and it is common in Cephalotaxus and most Podocarpaceae [5]. In Cupressaceae it is present in all genera.
When axial parenchyma cells are cut transversely, they are very similar to the adjacent axial tracheids, but have thinner walls and frequently include dark contents [93]. The arrangement of axial parenchyma in the transverse section can be used for identification. It must be observed at low magnification, viewing as many growth rings as possible because in some taxa with regular presence of axial parenchyma it can be scarce or absent in a single ring. Three patterns of arrangement can be distinguished (Figure 26). Diffuse: axial parenchyma cells single or in pairs distributed evenly along growth rings (Cephalotaxus and Podocarpaceae); tangentially zonate: parenchyma cells distributed in lines of different lengths parallel or oblique to the growth ring, occurring most frequently in the earlywood/latewood transition zone and in latewood, e.g., in Cupressaceae s.l. [19], conspicuous in the genera Callitris, Calocedrus, Chamaecyparis, Cryptomeria, Cupressus, Juniperus, Taiwania, Taxodium and Thuja [5]; and marginal: single axial parenchyma cells distributed along the growth ring boundary in the last row of latewood and/or first row of earlywood, e.g., in all genera of Abietoideae [14]. In addition, a single species can present all three arrangements.
The transverse end walls of axial parenchyma cells, observable in longitudinal sections, can be smooth, irregularly thickened, or nodular (Figure 27); in the latter case is generally more conspicuous in the tangential section. Although the morphology of the transverse end walls is used as a diagnostic feature in conifers, in Cupressaceae it must be interpreted with caution, as both smooth and irregular walls appear in some taxa, e.g., Glyptostrobus [83], Fitzroya [140], Sequoiadendron, Cryptomeria, Diselma and Fokienia [56], as well as Tetraclinis [116]. In Cupressaceae, moreover, the nodular appearance of transverse end walls is caused by the thickening of the primary wall rather than pitting in the strict sense, while in Abies, Cathaya, Keteleeria, Larix, Picea, Pseudotsuga and Tsuga, nodules have a similar appearance but exist due to the true pitting of the secondary wall [52]. In any case, this feature must be observed in mature wood, as its appearance can differ in juvenile wood.
End walls are smooth in Callitris, Callitropsis nootkatensis. (Syn.—Xanthocyparis nootkatensis (D.Don) Farjon and Harder), Tetraclinis articulata, Thuja occidentalis, Widdringtonia spp. (Cupressaceae), Dacrydium cupressinum and Podocarpus spp. (Podocarpaceae). Irregularly thickened walls occur in Chamaecyparis thyoides and Cryptomeria japonica. Markedly nodular walls are observed in Taxodium distichum, Calocedrus decurrens and C. formosana, Chamaecyparis obtusa and C. pisifera, some species of Juniperus, Thuja standishii, and Thujopsis dolabrata (Cupressaceae), and Abies, Cedrus, Keteleeria, Pseudolarix, Pseudotsuga and Tsuga (Pinaceae) [5].
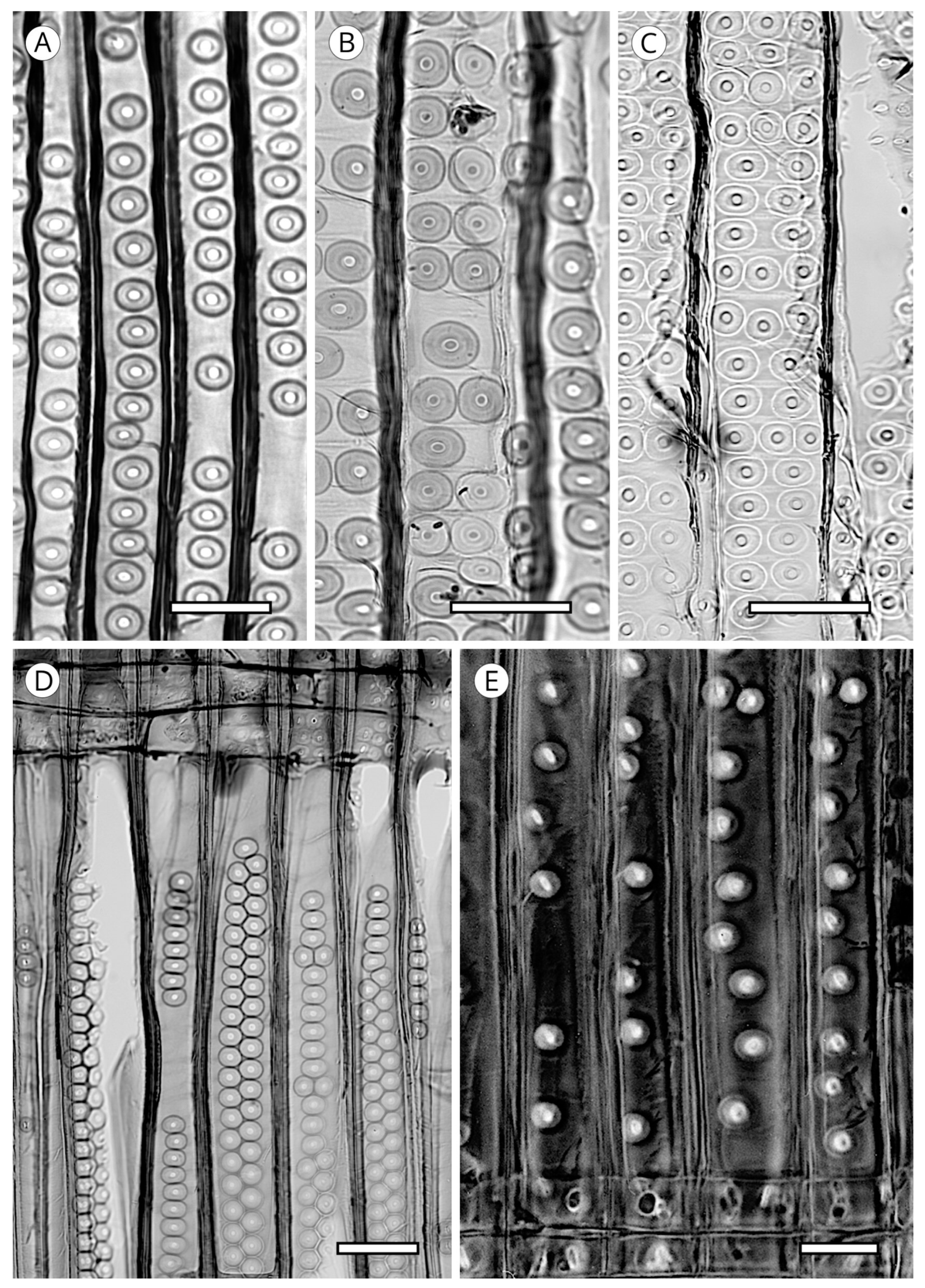
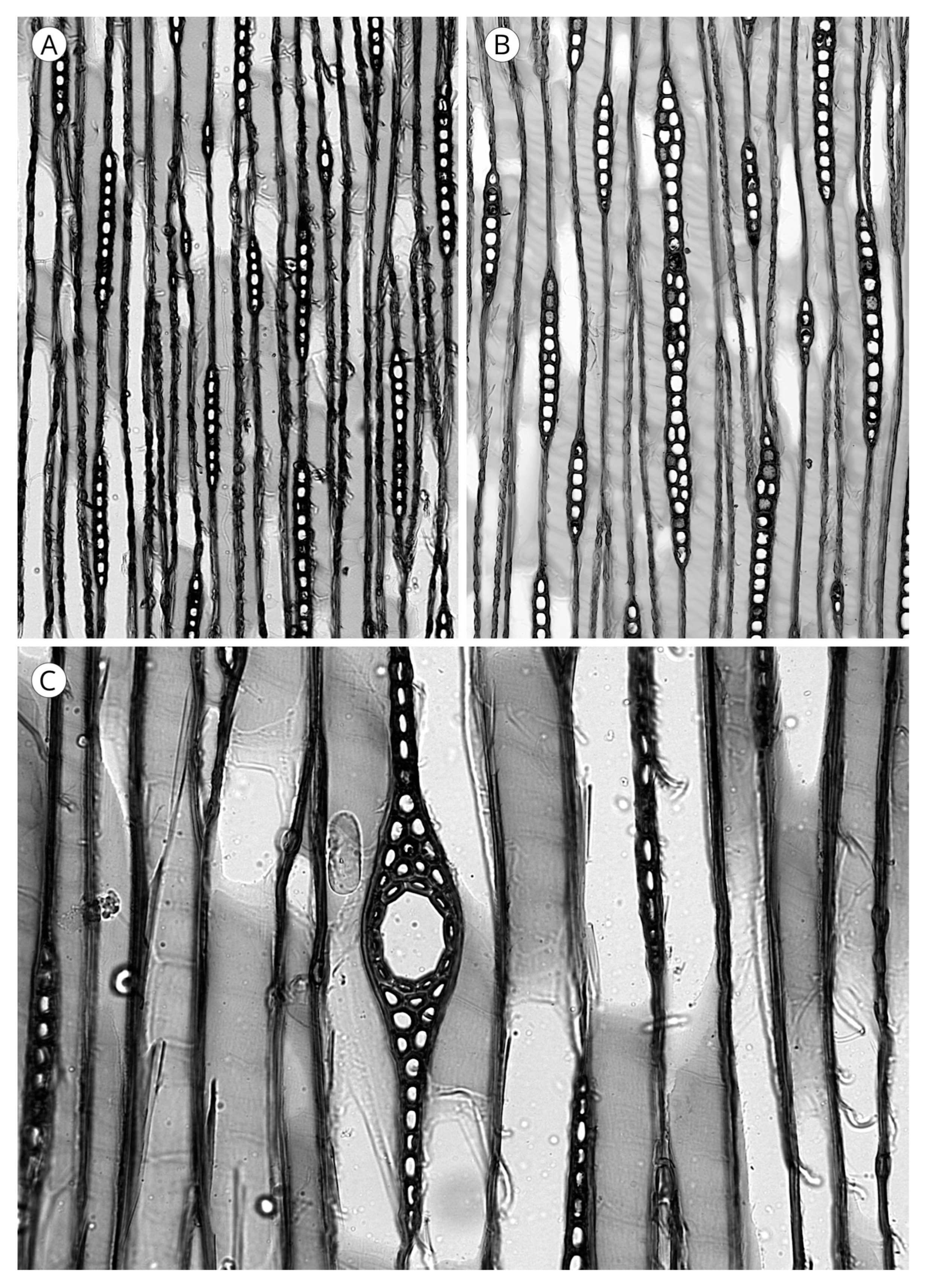
5.3. Rays
One of the unique wood features of today’s conifers is that their rays are uniseriate and occasionally biseriate in some species (Figure 28) [56]. However, the term biseriate is attributed to a sample only when at least 10% of all larger rays are biseriate over nearly the full height [5]. Only the presence of radial resin canals in taxa with longitudinal axial resin canals (Cathaya, Larix, Picea, Pinus and Pseudotsuga) causes the presence of thickened fusiform rays around the canal (Figure 28). In some species of the genus Pinus, particularly in section Strobus (e.g., P. cembra, P. koraiensis, P. lambertiana, P. monticola and P. strobus), the uniseriate extensions at both ends of the ray may be very high, and this must be taken into account when measuring ray height, ensuring the entire ray is included in the total measurement [5].
With regard to ray height, fossil records from the Carboniferous have demonstrated the coexistence of wood with uniseriate rays one or two cells high, as in the genus Bilignea, and wood with multiseriate rays up to 100 cells high in Calamopitys and Pitus [92,141,142].
Ray height is a good diagnostic feature in conifer wood. In Abies it is used, among other features, to separate the genus from other genera and even to differentiate groups of species in the genus [113,143]. Ray height is expressed in number of cells or in μm. Esteban et al. [56] and the IAWA Committee [5] proposed very similar thresholds for number of cells. The first authors proposed three categories (1 to 15, 16 to 30 and more than 30), while the IAWA Committee [5] proposed a fourth range in the lowest heights: up to 4 cells, 5 to 15, 16 to 30 and more than 30. In any case, ray height in number of cells can be a feature common to a group of softwoods in taxonomic terms, e.g., in Cupressaceae [93].
Ray height must always be measured in mature wood because of the differences between mature and juvenile wood, as it is lower in juvenile wood.
5.3.1. Ray Tracheids
A softwood ray in its most complex composition is made up of ray tracheids, ray parenchyma cells and resin canal epithelial cells. When it is composed of ray tracheids and parenchyma cells, it is termed heterocellular. In contrast, when it comprises only ray parenchyma cells, it is termed homocellular [94] (Figure 29). Although the position of tracheids in the ray has no diagnostic value, they may be above or below the ray, i.e., in marginal rows, e.g., Larix, Picea and Pinus, or in alternate rows, both marginal and inside the ray [64] (Figure 29). In some taxa, rays can be composed solely of tracheids, as in species of southern yellow pine and very rarely in species of Picea and Larix [65,131]. Ray tracheids have the same characteristics as axial tracheids, but differ slightly in morphology and pit size. Ray tracheids are usually more irregularly shaped, with less tapered end walls, especially in marginal arrangement, and ray tracheid pits are always smaller in diameter than pits in the radial walls of longitudinal tracheids [56].
In Pinaceae, ray tracheids are found only in taxa with normal resin canals, except for Tsuga, which lacks resin canals.
In Abietoideae, Tsuga invariably has ray tracheids, Cedrus frequently has them, Abies, Nothotsuga and Pseudolarix rarely have them and they are absent in Keteleeria. The absence of ray tracheids in Keteleeria has diagnostic value because it separates it from Nothotsuga, as they are the only two genera of all the conifers that have only normal axial resin canals [14].
Other non-Pinaceae species have ray tracheids, particularly Cupressaceae, e.g., Cupressus arizonica, Sequoia and Thujopsis dolabrata [52], and Callitropsis nootkatensis, which has some rays formed solely of tracheids and others solely of parenchyma cells [5] (Figure 29).
Hudson [144] proposed a classification of ray tracheids in Pinus with 14 groups based on degree of dentation, from very slight horizontal wall thickenings in P. cembra to reticulate in P. ponderosa, taeda and palustris.
The IAWA Committee [5] and Heinz [57] established three types of degree of dentation in ray tracheids: smooth, e.g., the “soft pines” group (Pinus, section Strobus: e.g., P. cembra, P. koraiensis, P. lambertiana, P. monticola and P. strobus); dentate, with dentations of variable thickness arranged as protrusions from the upper and lower cell wall of ray tracheids, very prominent in Pinus, e.g., sections Sylvestris (e.g., P. densiflora, P. nigra, P. resinosa and P. sylvestris) and Ponderosa (e.g., P. contorta, P. patula, P. pinaster, P. ponderosa and P. radiata); less prominent and with typically sinuous tracheid walls, e.g., in Pinus, sections Sula (P. canariensis, P. halepensis, P. leucodermis, P. longifolia) and Khasya (P. khasya); inconspicuous, as in some species of Picea [52]; and reticulate, when thickenings are thin-walled, very numerous, narrow, with tooth-like protrusions from the top to the bottom of the tracheid, resulting in a characteristic reticulate appearance, e.g., Pinus, section Taeda (including, i.a., P. banksiana, P. palustris, and P. taeda) (Figure 30). Some authors [119,145] studied the morphology of tracheid dentations in an attempt to differentiate very closely related species, e.g., Pinus nigra and P. sylvestris, but the results obtained were not conclusive.
Esteban et al. [129], following the classification of the three types of dentations, included the following species of Pinus: smooth (P. armandii, P. ayacahuite, P. brutia, P. cembra, P. cembroides, P. gerardiana, P. griffithii, P. halepensis, P. hartwegii, P. koraiensis, P. merkusii, P. monophylla, P. morrisonicola, P. parviflora, P. peuce, P. pinea, P. quadrifolia, P. strobus and P. wallichiana), dentate (P. arizonica, P. banksiana, P. brutia, P. canariensis, P. caribaea, P. cembroides, P. contorta, P. densiflora, P. devoniana, P. douglasiana, P. elliottii, P. gerardiana, P. griffithii, P. halepensis, P. hartwegii, P. heldreichii, P. jeffreyi, P. kesiya, P. lawsonii, P. leiophylla, P. massoniana, P. maximinoi, P. merkusii, P. monophylla, P. montezumae, P. mugo, P. nigra, P. palustris, P. pinaster, P. pinea, P. ponderosa, P. pseudostrobus, P. quadrifolia, P. radiata, P. resinosa, P. rigida, P. sylvestris, P. tabuliformis, P. taeda, P. tecunumanii, P. teocote, P. thunbergii and P. tropicalis), and reticulate (P. arizonica, P. banksiana, P. caribaea, P. contorta, P. devoniana, P. douglasiana, P. elliottii, P. heldreichii, P. jeffreyi, P. maximinoi, P. palustris, P. ponderosa, P. rigida, P. taeda and P. tecunumanii).
In the ray tracheids of some species, helical thickenings develop with the same characteristics as thickenings in axial tracheids (Figure 30), e.g., Cathaya argyrophylla [128], Pseudotsuga sinensis [129], some species of Larix (L. potaninii var. himalaica [69] L. sibirica [57], and Picea (P. polita) [60], P. spinulosa and P. torano [129]. In other species, e.g., Pseudotsuga menziesii and Larix kaempferi, helical thickenings rarely occur [5] and have no diagnostic value due to their variable occurrence [57].
In the radial section, Bartholin [66] studied the morphology of the aperture in ray tracheid pit borders in Picea, distinguishing two types: pits that are thickened by small lumps, giving the aperture the appearance of a narrow canal (Picea-1 type), and thickenings resembling small horns on the edge of the pit aperture (Picea-2 type). This anatomical peculiarity enables differentiation between Picea and Larix, as the latter has ray tracheid pits with larger apertures [66,130] (Figure 31).
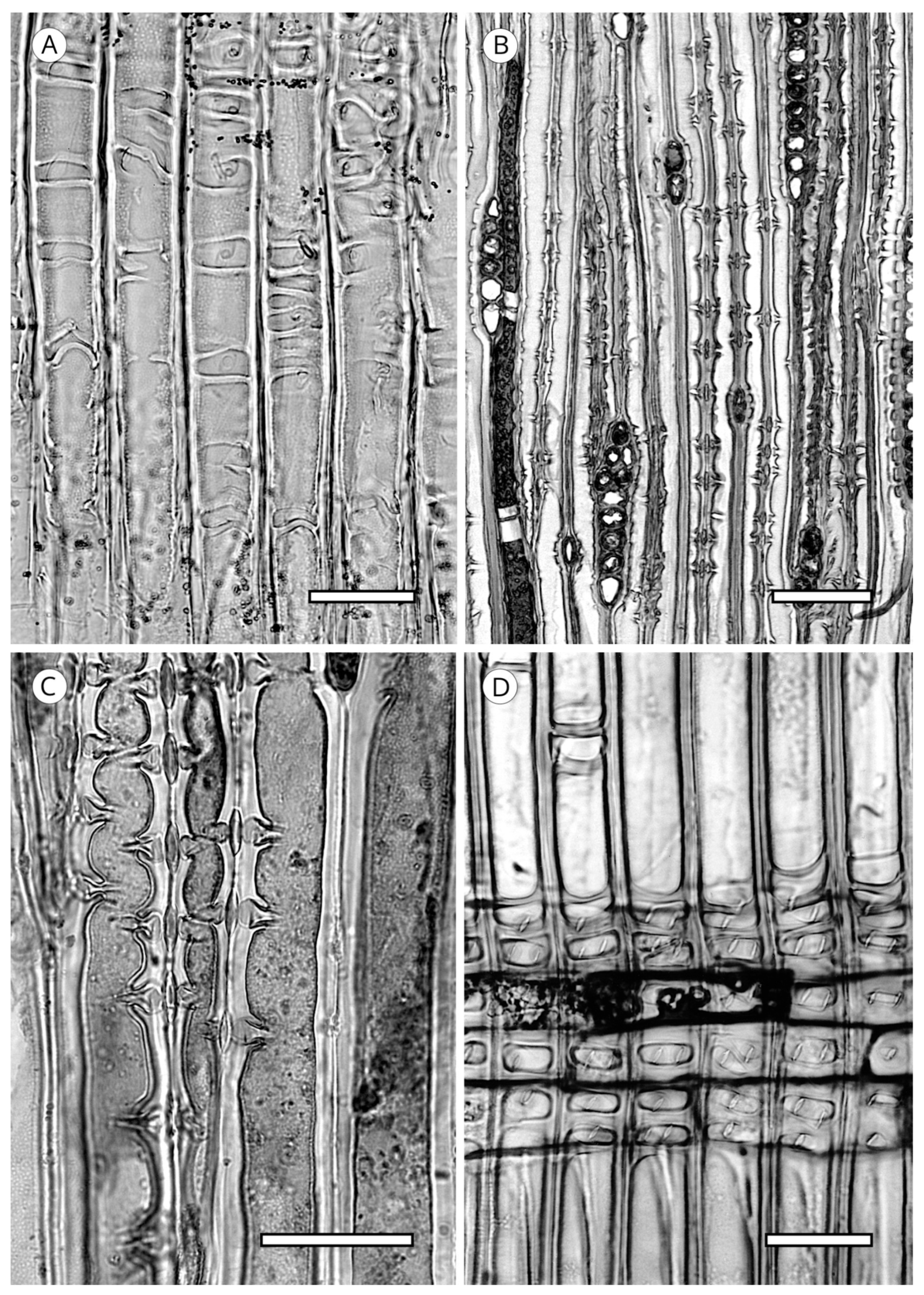
5.3.2. Ray Parenchyma
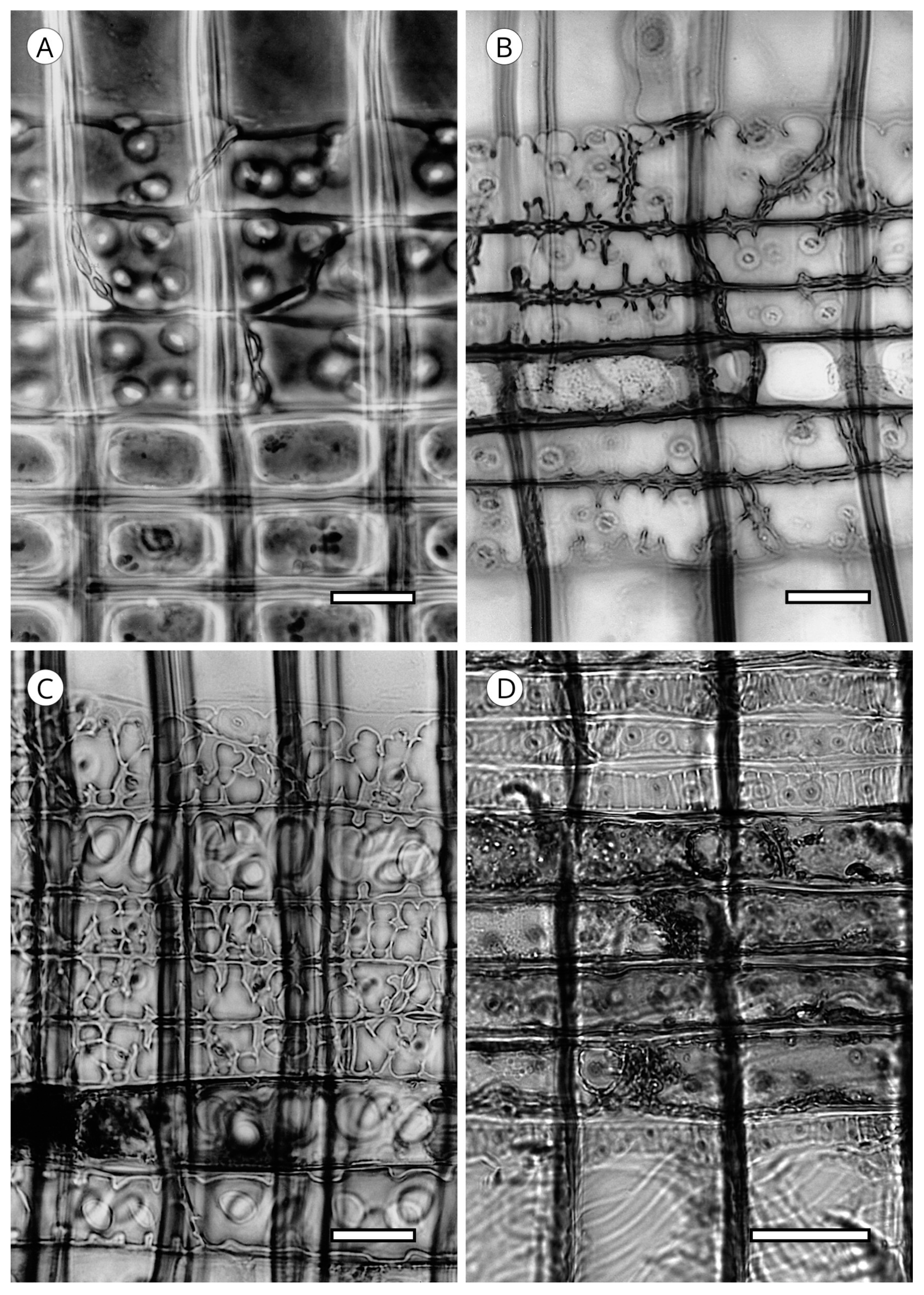
Most conifer wood has horizontal walls of ray parenchyma cells that are thin and have no pitting. Species with pitted horizontal walls are limited to all genera of Pinaceae (Abies (Figure 32B,C), Cathaya, Cedrus, Keteleeria, Larix, Nothotsuga, Picea, Pseudotsuga and Tsuga), although numerous species of Pinus do not conform to this [5,57,129]; pitted horizontal walls are inconspicuous in Pseudolarix [14]. Heinz [57] described intermediary situations with the presence of smooth to slightly pitted horizontal walls in Calocedrus, Cryptomeria, Cupressus, Juniperus, Metasequoia, Platycladus, Taxodium (Figure 32A), Thuja (Cupressaceae), Taxus and Torreya (Taxaceae).
Like the horizontal walls, end walls in most conifer woods are thin, with little or no pitting. Nodular end walls are limited to the genera of Pinaceae (absent in some species of Pinus, slightly nodular in Pinus section Strobus (e.g., P. cembra, P. koraiensis, P. lambertiana, P. monticola and P. strobus) [5] and Cupressaceae. In Abies they are present in all species [113]. They are considered characteristic of this genus [52,53,146] and can be used to differentiate Abies from the other conifer genera, except for those in Abietoideae, as the presence of nodules in end walls is common to the whole subfamily [14]. In Cupressaceae, nodular end walls are different in appearance, occurring in Calocedrus, Cunninghamia (normally smooth), Cupressus, Diselma, Fitzroya, Juniperus and Xanthocyparis [86] (Figure 33). From our observations in the two species of Cunninghamia we cannot confirm that they are mostly smooth. A review of the literature shows that most authors report smooth end walls, whereas we observed both smooth and distinctly nodular end walls. Moreover, some genera, especially Juniperus and Cupressus, have smooth and nodular end walls in a single species and many transitional forms can be found, and therefore, this feature should be used with caution.
At the junction of end walls with horizontal walls of ray parenchyma cells, depressions can appear. Peirce [47] first named them indentures, reporting them as particularly common in the genera Cunninghamia and Taiwania (Cupressaceae). According to Phillips [52], indentures occur in all conifer families except Araucariaceae; in Podocarpaceae they are present only in Podocarpus salignus and Dacrycarpus (Podocarpus dacrydioides), and in Cedrus, Keteleeria and Pinus (Pinaceae) they are only poorly developed. However, Esteban and de Palacios [14] reported that they are clearly visible in all genera of Abietoideae (Abies, Cedrus, Keteleeria, Nothotsuga, Pseudolarix and Tsuga). Yatsenko-Khmelevsky [131] stated that in Cedrus, Keteleeria and Pinus they are poorly developed or absent, while according to Farjon [19], they must be considered a feature of diagnostic value in Cupressaceae s.l. In fact, Román-Jordán [86] recorded them in Austrocedrus, Calocedrus, Cryptomeria, Cunninghamia, Fokienia, Glyptostrobus, Pilgerodendron, Sequoia, Taxodium and Xanthocyparis (Figure 34). To avoid misidentification, this feature should be used with caution and not in wood with poorly defined and rare indentures [5].
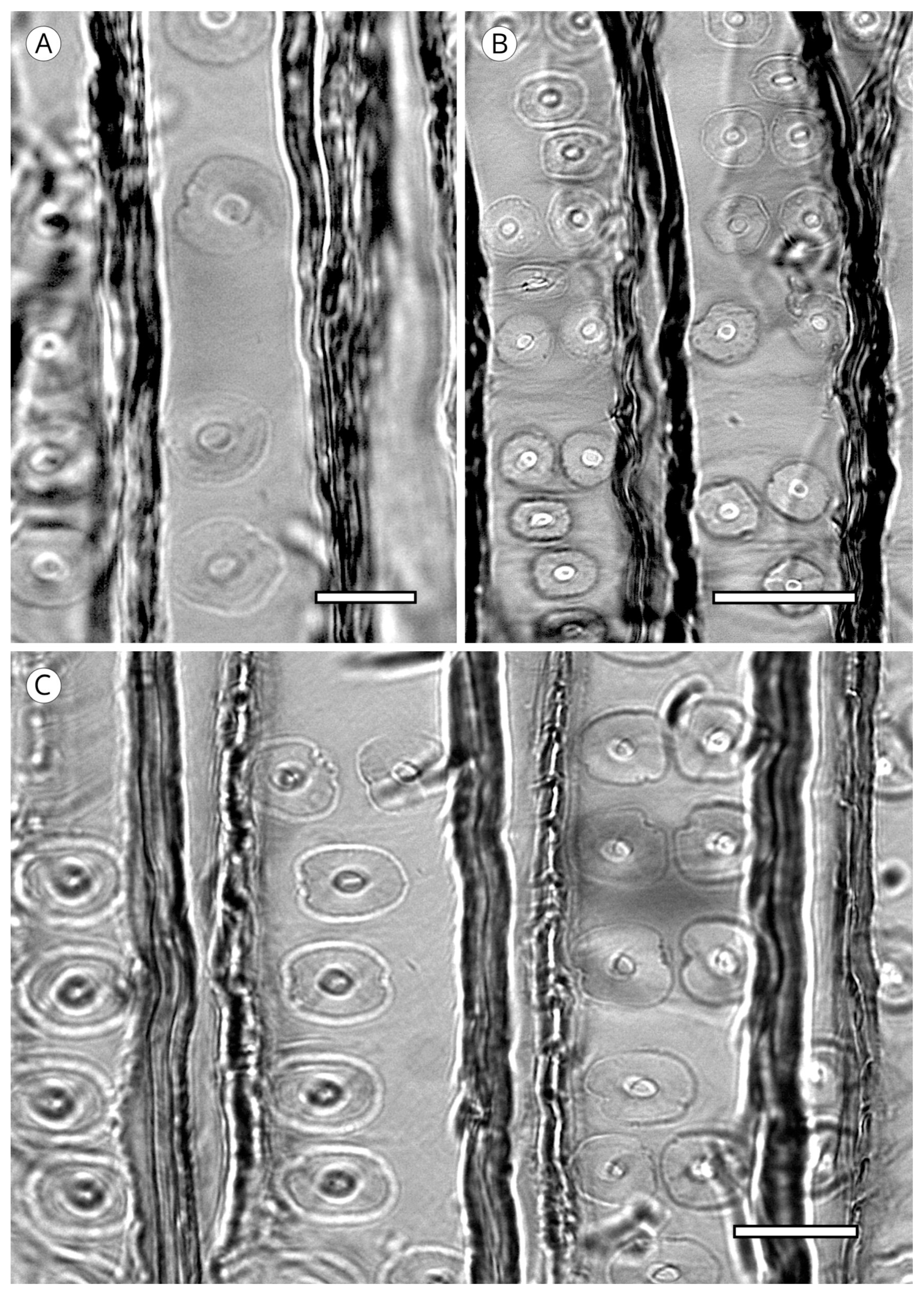
5.3.3. Cross-Field Pitting
Cross-field pitting is undoubtedly one of the most valuable analytical features for softwood identification. Pits are located in the area of contact between the intersecting walls of a single longitudinal tracheid and a single ray parenchyma cell as seen in radial section. The cross-field pit features analysed are the frequency, arrangement, form, size and/or position of the aperture relative to the border of the pits.
Cross-field pit types are based on the classification established by Phillips [52] to which the IAWA Committee [5] added the araucarioid type used by Barefoot and Hankins [55] and re-evaluated by Vogel [73] (Figure 36 and Figure 37):
Window-like. Pits occupy nearly the entire cross-field. They are square or rectangular and there are usually one or two per cross-field. They have almost no borders and are exclusive to the genus Pinus (sections Strobus and Sylvestris) (Pinaceae) IAWA Committee [5], Lagarostrobos, Phyllocladus, Sundacarpus amarus (Podocarpaceae) and Sciadopitys (Sciadopityaceae) (Phillips, 1941) [52]. According to Rol [147], Pinus kesiya and P. merkusii should be included as examples of taxa that present these types of pits with more than two pits per cross-field [5].
Pinoid. These pits are simple or have almost no border. They are irregularly shaped and highly variable in number, with up to six pits per field, normally with three or more. Pinoid pits are exclusive to the species of the genus Pinus that do not have window-like pits.
Piceoid. Pits have borders and the apertures, in this case, are long, narrow, typically oval and extend beyond the borders. These are the smallest of all the pit types. This type of pitting occurs in Pinaceae in Cathaya [128], Larix, Picea and Pseudotsuga [5] and all the genera of Abietoideae except Nothotsuga [14].
Cupressoid. Pits have borders and the oval apertures are contained entirely within them. This type of cross-field pitting is characteristic of Cupressaceae, except for the genus Thuja, where pitting is taxodioid. It is also present in Podocarpaceae and Taxaceae.
Taxodioid. The are large pits, oval to circular, with apertures of the same shape. The aperture exceeds the width of the border at its widest point. These pits occur in most of the former family Taxodiaceae (now Cupressaceae), e.g., Cunninghamia, Sequoia, and Taxodium, although they have also been reported in Thuja (Cupressaceae), in some species of Podocarpaceae [5] and in Abietoideae (Abies, Cedrus, Nothotsuga and Pseudolarix) [14]. However, taxodioid pits in the subfamily Abietoideae are considerably smaller and similar in size to piceoid pits, with included apertures within the limits of the pit border (contrary to the often extended piceoid pits). In the literature they are also referred to as “abietoid” pits [53]. In some Cupressaceae genera, in particular Sequoia and Taxodium, pits are usually arranged in rows of two or three per cross-field except in marginal cells [5]. Many transitional forms between cupressoid and taxodioid cross-field pitting occur, and therefore, both types should be taken into account for identification if necessary.
Araucarioid. Restricted to Araucariaceae (Agathis, Araucaria and Wollemia). These are cupressoid pits but with a different grouping, with a tendency to crowd, which gives pit perimeters the appearance of alternate polygonal pits (honeycomb-like) similar to those in axial tracheids in Araucariaceae.
5.4. Resin Canals
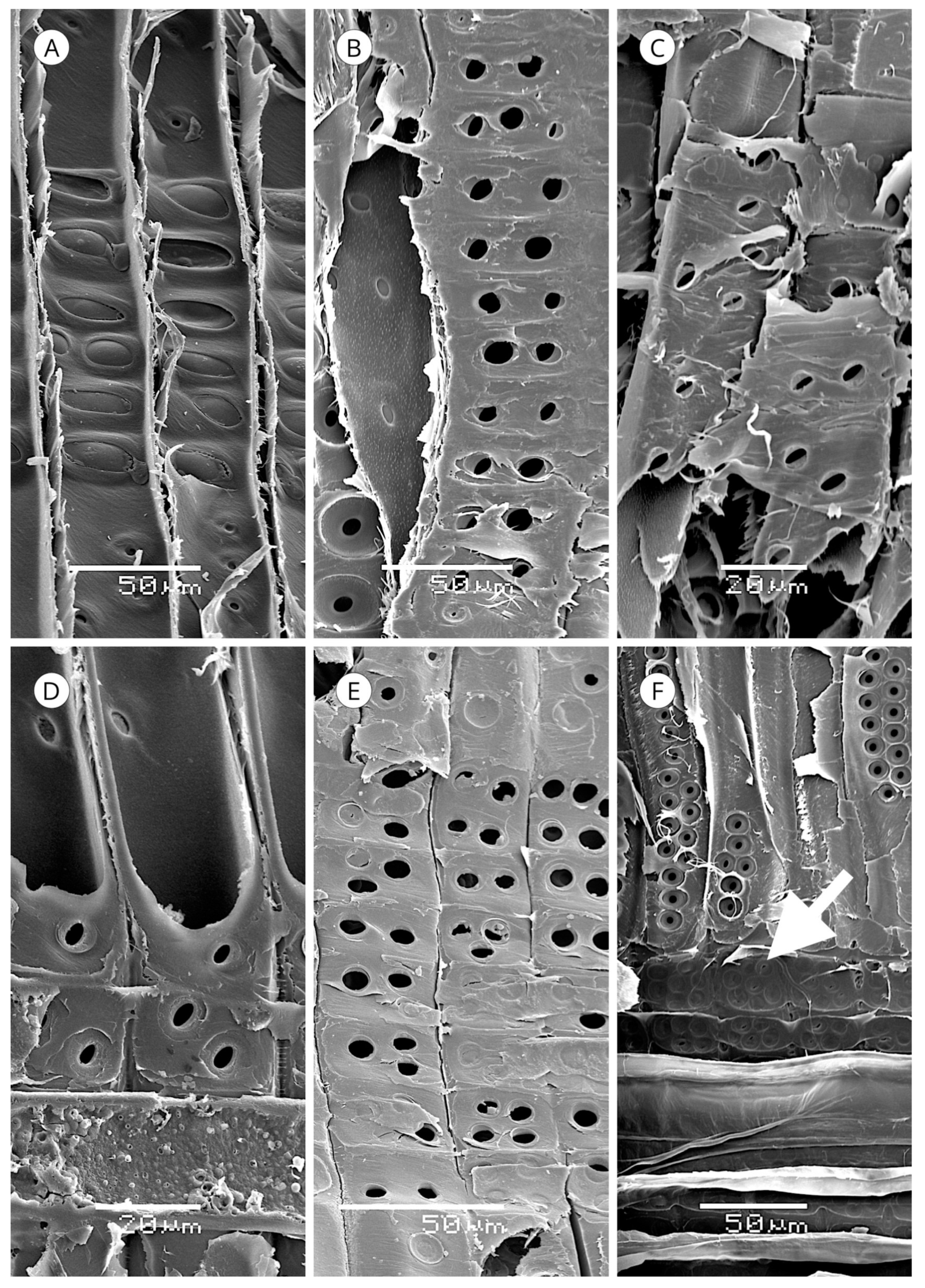
Resin canals are postcambial tubular structures formed by the separation of groups of contiguous cells (schizogenesis) [93]. The cells lining the tubular cavity are derived from fusiform cells in the cambium that have undergone mitotic divisions [67,148,149] from the same type of cells that form axial tracheids. However, rather than maturing into tracheids, they remain as parenchyma cells, producing transverse walls [93]. The cells enclosing the whole tube constitute the epithelium of the sheath of the resin canal.
In Pinaceae, some taxa have subsidiary cells around the canals (e.g., Pinus canariensis) [120], and although some authors called them axial parenchyma cells (e.g., Kibblewhite and Thompson [148] in Pinus elliottii, LaPasha and Wheeler [70] in Pinus taeda, Howard and Manwiller [134] in the southern yellow pines, Fahn [150] in Pinus densiflora, Ickert-Bond [151] in Pinus krempfii), Wiedenhoeft and Miller [82] unified the terminology of resin canals, defining the canal as the intercellular space itself and the epithelium as the single layer of cells lining the canal, reserving the term subsidiary cells for the cells outside the epithelium, which can be subsidiary parenchyma and/or strand tracheids. Unlike normal axial tracheids, strand tracheids are shorter and divided by transverse walls containing normal tracheid pits. Some authors considered them to be transitional elements between axial tracheids and epithelial cells or axial parenchyma cells [93], although they based this solely on the association of strand tracheids with one cell type or another (Figure 38).
In Cathaya, Larix, Picea and Pseudotsuga, the resin canal is composed solely of the canal itself and the epithelium. However, some authors have reported subsidiary cells in some species of these genera, e.g., Takahara et al. [152] in Picea abies and Sato and Ishida [153] in Larix kaempferi. In Pinus the presence of more than one layer of cells is very frequent [82].
Radial resin canals have similar ontogeny. The canal cavity is also formed by schizogenesis, but in this case, cells with transverse walls are not divided from long cells into shorter units. Some ray cells, rather than becoming ray parenchyma or ray tracheids, are directly converted into epithelial cells [93].
Epithelial cells, which can be thin- or thick-walled, are where the resin released into the canal is produced.
Resin canals are a key feature for the taxonomic position of conifers [44,53,123,150,154,155,156]. The presence of normal resin canals in Pinaceae has led to various classifications of the family.
Jeffrey [157] made two divisions in Pinaceae: the presence of resin canals in seeds supported the monophyly of Abies, Cedrus, Keteleeria, Pseudolarix, Tsuga, and resin canals in secondary xylem and leaves with endodermis having thickened Casparian strips supported the monophyly of Cathaya, Larix, Picea, Pinus and Pseudotsuga. Wu and Hu [155] divided Pinaceae into three groups: with normal resin canals (Cathaya, Pinus, Larix, Pseudotsuga and Picea); without normal resin canals (Abies, Pseudolarix, Cedrus and Tsuga); and normal resin canals absent or present in Keteleeria. After confirming the permanent presence of axial resin canals in mature wood in all species of Keteleeria and the pattern of traumatic resin canals, Lin et al. [156] proposed a classification of Pinaceae in three groups: with radial and axial resin canals (Cathaya, Larix, Picea, Pinus and Pseudotsuga); with only axial resin canals (Keteleeria and Nothotsuga) and without resin canals (Abies, Pseudolarix, Cedrus and Tsuga). Esteban and de Palacios [14] confirmed this classification, based on the resin canals and other features of the secondary xylem. Lin et al. [158] suggested that on the basis of the presence of axial resin canals in latewood and resin cavities in their bark, Nothotsuga and Keteleeria have a high number of similarities. Therefore, based on the presence of resin canals, Abietoideae supports two clearly distinct groups: Abies, Cedrus, Pseudolarix and Tsuga without resin canals, and Keteleeria and Nothotsuga with only axial resin canals.
Various authors have classified the anatomy of resin canals based on the presence of thick- or thin-walled epithelial cells [64,146,149,159,160,161,162], while others have attempted to establish the number of epithelial cells surrounding the canals in taxa with thick-walled cells as a distinguishing feature [55,56,60,63]. There is definitely some discrepancy in the intervals proposed, and therefore, the IAWA Committee [5] did not include this feature. Moreover, the IAWA Committee [5] indicated that at times it is difficult to establish a division between thin- and thick-walled epithelial cells, e.g., in Larix and Picea, because the two types can be mixed. Nonetheless, overall, two large groups of softwood with resin canals can be distinguished based on the epithelial cells: thick-walled (Cathaya, Keteleeria, Larix, Nothotsuga, Picea and Pseudotsuga) and thin-walled (Pinus).
Both axial and radial resin canals can be normal or of traumatic origin [94]. Normal resin canals occur only in some genera of Pinaceae. Cathaya, Larix, Picea, Pinus and Pseudotsuga have axial and radial resin canals, while Keteleeria [52,79,146,156,163] and Nothotsuga [158] have only axial resin canals. The absence of resin canals in some descriptions of Nothotsuga [154,155,157] is probably because the wood analysed was juvenile or from branches. No conifer taxon contains only normal radial resin canals (Figure 39 and Figure 40).
Traumatic resin canals are formed in response to wounding in Pinaceae, both in genera with normal resin canals (Cathaya, Keteleeria, Larix, Nothotsuga, Picea, Pinus and Pseudotsuga) and in genera without them (Abies, Cedrus, Nothotsuga, Pseudolarix and Tsuga) (Figure 41). In non-Pinaceae genera, they have been reported in Sequoia sempervirens [164], Sequoiadendron [25], Microbiota [165] and Tetraclinis [116].
In Abies [146,166,167,168], Tsuga [5] and Pseudolarix [155], only axial traumatic resin canals occur, while in Cedrus, both axial and radial traumatic canals can be present [14,169]. This is the only genus without normal resin canals in which both types of traumatic resin canals occur simultaneously. Cedrus is, therefore, the only genus of Pinaceae (Abietoideae) capable of forming both axial and radial traumatic resin canals as a response to wounding, and consequently, some authors have considered it an intermediary stage between Pinus and Abies [167]. According to Gernandt et al. [15], molecular evidence supports the genus Cedrus as a sister group to the other Pinaceae and should occupy a basic or root position in the family, such that the subfamily Abietoideae can be considered paraphyletic.
Some authors reported the presence of normal resin canals in Abies, e.g., Penhallow [170] in A. concolor, A. bracteata, A. nobilis and A. firma and Vierhapfer [171] in A. concolor, A. nobilis and A. bracteata, but Abies, Cedrus, Tsuga and Pseudolarix lack normal resin canals and form canals only in response to wounding. The four genera have features characteristic of traumatic resin canals, with an irregular perimeter and all appearing in tangential rows.
Although traumatic resin canals were not observed in Pseudolarix, in this study, they were reported in the genus as a response to wounding by Wu and Hu [155].
With regard to wood that has only normal axial resin canals, both Keteleeria and Nothotsuga form traumatic axial resin canals in response to wounding, distributed randomly in single arrangement and/or tangential rows [156]. This arrangement differs from the arrangement in traumatic resin canals in Abies, Tsuga, Pseudolarix [44,167] and Cedrus [155], where they are invariably in tangential rows.
For resin canal biometry, one of the main problems in reviewing the literature with regard to size is the lack of indications about the method used. This makes it difficult to compare the results obtained by different authors. The three methods established by the IAWA Committee [5] will undoubtedly solve this problem.
Method A. The tangential diameter of the resin canal, including the epithelial cells, is measured at the widest part of the opening. This is the most frequently used method.
Method B. The tangential diameter of the entire resin canal complex is measured at the widest point, taking in all components of the axial resin canal to the point of differentiation from the axial tracheids.
Method C. The radial diameter of the whole canal complex, including the epithelial cells, is measured at the widest point.
Some authors [53,64,67,93,161] considered that the resin canal size can be used as a diagnostic feature. Others, however, [52,54,172] did not consider it a diagnostic method.
This divergence of criteria is probably due to which of these three methods explained by the IAWA Committee [5] is used, as the choice of method could depend on the species. For example, Wiedenhoeft et al. [173] considered that method A [5] does not allow separation of Pinus contorta from P. ponderosa, whereas method B, which measures the tangential width of the entire resin canal complex [5], is more reliable.
5.5. Mineral Inclusions
The presence of calcium oxalate crystals is not very common in coniferous woods (Figure 42). They have been reported in several species of Abies [59,113] in all the genera of Abietoideae [14], in Picea, in the form of prismatic crystals, and in Pinus flexilis as small styloid crystals. Their regular presence has considerable diagnostic value [5]. In addition, crystals in conifers appear to be located in a single cell type in a given taxon. For example, crystals are arranged in marginal and submarginal rows of ray parenchyma in all genera of Abietoideae [14] and in some species of Picea [5]. They have also been observed in the epithelium of axial and radial resin canals in the form of styloids in Pinus bungeana [57], Pinus flexilis [77], Pinus aristata, P. balfouriana, P. longaeva [68] and Pinus contorta [173]. Organic crystalline deposits have been reported in axial tracheids in Tsuga heterophylla [174], Callitris endlicheri [74] and Torreya yunnanensis [75].
6. Families and Genera
Araucariaceae Henkel and W.Hochst.—three genera: Araucaria Juss. (20 species), Agathis Salisb. (17), Wollemia W.G.Jones, K.D.Hill and J.M.Allen (1). Axial tracheids—intercellular spaces occasional. Tracheid pits in radial walls uniseriate and/or biseriate, also multiseriate, all polygonal in appearance; biseriate/multiseriate pits in alternate arrangement. Torus defined. Axial tracheids with organic content, often forming groups close to the rays. Additionally, in spools, trabeculae-like. Helical thickenings absent. Callitroid thickenings absent. Axial parenchyma—rare or absent. Rays— ray tracheids absent. Smooth horizontal and end walls. Cross-field pitting araucarioid. Resin canals— absent. Mineral inclusions—not observed.
Cupressaceae Gray—thirty two genera: Actinostrobus Miq. (3 species), Athrotaxis D.Don (3), Austrocedrus Florin and Boutelje (1), Callitris Vent. (20), Callitropsis Oerst. (1), Calocedrus Kurz (4), Chamaecyparis Spach (5), Cryptomeria D.Don (1), Cunninghamia R.Br. (2), Cupressus L. (14), Diselma Hook.f. (1), Fitzroya Lindl. (1), Fokienia A.Henry and H.H.Thomas (1), Glyptostrobus Endl. (1), Hesperocyparis Bartel and R.A.Price (17), Juniperus L. (68), Libocedrus Endl. (5), Metasequoia Hu and W.C.Cheng (1), Microbiota Kom. (1), Neocallitropsis Florin (1), Papuacedrus H.L.Li (1), Pilgerodendron Florin (1), Platycladus Spach (1), Sequoia Endl. (1), Sequoiadendron J.Buchholz (1), Taiwania Hayata (1), Taxodium Rich. (1), Tetraclinis Mast. (1), Thuja L. (5), Thujopsis Siebold and Zucc. ex Endl. (1), Widdringtonia Endl. (4), Xanthocyparis Farjon and T.H.Nguyên (1). Axial tracheids—frequent presence of intercellular spaces in Cupressus and Juniperus. We also observed them in Calocedrus formosana, Chamaecyparis lawsoniana, Pilgerodendron and some species of Callitris (C. columellaris, C. endlicheri and C. neocaledonica). Tracheid pitting in radial walls uniseriate and/or biseriate. Biseriate normally in Sequoia and Sequoiadendron, up to triseriate in Taxodium. Torus defined, except in some species of Thuja and Thujopsis. Torus extensions were present in Actinostrobus, Athrotaxis, Austrocedrus, Cupressus dupreziana, C. funebris, Diselma, Fitzroya, Juniperus brevifolia, J. californica, J. deppeana, J. monosperma, J. monticola, J. osteosperma, J. pinchotii, J. scopulorum, J. standleyi, Pilgerodendron and Widdringtonia. Pits with notched borders were present in Athrotaxis, Calocedrus decurrens, Chamaecyparis pisifera, Cryptomeria, Cunninghamia, Cupressus dupreziana, Juniperus brevifolia, J. thurifera, Papuacedrus, Sequoia, Taiwania, Thuja occidentalis, Thujopsis and Callitropsis nootkatensis. This feature is used to differentiate Sequoia (notched borders present) from Sequoiadendron (notched borders absent) [101]. It was also used by de Palacios et al. [85] to differentiate Macaronesian species of Juniperus. Habitual presence of warty layer in the family, although we did not observe it in Libocedrus, Neocallitropsis or Papuacedrus, less conspicuous in Athrotaxis, Austrocedrus, Calocedrus, Cryptomeria, Cunninghamia, Fokienia, Metasequoia, Sciadopitys, Sequoia, Sequoiadendron, Taiwania, Thujopsis, Xanthocyparis and some species of Chamaecyparis and Juniperus. Very clear in Callitris, Actinostrobus, Tetraclinis and some species of Juniperus. Organic deposits regularly occur in some Cupressaceae genera. They have been reported in Callitris columellaris, Calocedrus decurrens, Cupressus, Fitzroya, Juniperus procera, J. virginiana, Libocedrus bidwillii, Pilgerodendron, Thuja plicata and T. standishii [55,175,176]. We observed them in Widdringtonia and some species of Callitris (C. endlicheri, C. preissii and C. rhomboidea). Helical thickenings were absent. Callitroid thickenings were present in Actinostrobus and Callitris, occasionally in Neocallitropsis. Axial parenchyma— regularly present in all genera. Simultaneous occurrence of arrangements (diffuse, tangential and marginal) is common, e.g., in Actinostrobus pyramidalis, Calocedrus decurrens, C. macrolepis, Chamaecyparis pisifera, Hesperocyparis sargentii (Syn.—Cupressus sargentii Jeps.), Cupressus sempervirens, Glyptostrobus pensilis, Juniperus barbadensis, J. chinensis, J. deppeana, J. excelsa, J. oxycedrus and Tetraclinis articulata). Transverse end walls smooth, irregular or nodular or even transitional; also simultaneous occurrence of types, particularly in Cupressus and Juniperus. We observed smooth walls or smooth and occasionally irregular walls in Actinostrobus, Athrotaxis, Austrocedrus, Callitris and Cunninghamia, most Cupressus, Juniperus bermudiana, J. oxycedrus, J. procera, J. standleyi, J. taxifolia, Libocedrus, Metasequoia, Microbiota, Papuacedrus, Taiwania, Tetraclinis and Widdringtonia. Transverse end walls nodular or occasionally irregular and nodular in Calocedrus, Hesperocyparis bakeri (Syn.—Cupressus bakeri Jeps.), H. macnabiana (Syn.—Cupressus macnabiana A.Murray bis), Glyptostrobus, most Juniperus, Platycladus, Taxodium distichum and Thujopsis. Simultaneous occurrence of smooth, irregular and nodular transverse end walls in Chamaecyparis, Cryptomeria, Hesperocyparis goveniana (Syn.—Cupressus goveniana Gordon), H. macrocarpa (Syn.—Cupressus macrocarpa Hartw.), Diselma, Fokienia, some species of Juniperus, Pilgerodendron, Sequoia and Sequoiadendron. Very few genera have exclusively nodular transverse end walls: Calocedrus, Platycladus and Thujopsis. Rays—regular presence of ray tracheids in Callitropsis nootkatensis. Horizontal walls of ray parenchyma cells smooth (e.g., Diselma and Fitzroya), at times lightly pitted. End walls smooth and/or nodular, or even transitional forms. Cross-field pitting cupressoid (Cupressus and Juniperus), taxodioid (Athrotaxis, Cryptomeria, Cunninghamia, Fokienia, Metasequoia, Papuacedrus, Sequoia and Sequoiadendron), at times cupressoid and partially taxodioid (e.g., Austrocedrus, Calocedrus, Chamaecyparis, Fitzroya, Libocedrus, Pilgerodendron, Taiwania, Taxodium, Thuja, Thujopsis and Xanthocyparis). Resin canals—absent. In the samples studied, traumatic axial resin canals were observed in Diselma, Sequoia and Tetraclinis. They have also been described in Actinostrobus pyramidalis [29], Callitris drummondii [126], Chamaecyparis obtusa [177], Cunninghamia lanceolata [178], Cupressus funebris [179], Cryptomeria [180], Hesperocyparis arizonica (Syn.—Cupressus arizonica Greene) [170], Metasequoia [181], Microbiota [165], Sequoia [57] and Sequoiadendron [55]. Mineral inclusions—not observed.
Pinaceae Spreng. ex F.Rudolphi.—eleven genera: Abies Mill. (47 species), Cathaya Chun and Kuang (1), Cedrus Trew (3), Keteleeria Carrière (3), Larix Mill. (10), Nothotsuga H.H.Hu ex C.N.Page (1), Picea A.Dietr. (37), Pinus L. (118), Pseudolarix Gordon and Glend. (1), Pseudotsuga Carrière (4), Tsuga (Endl.) Carrière (10). Axial tracheids— tracheid pitting in radial walls uniseriate and/or biseriate, normally in opposite arrangement. Torus defined. Scalloped tori in Cedrus. Regular torus extensions in Abies and Tsuga. Regular presence of warty layer in Abies, Cedrus and Tsuga. Occasional organic deposits in Abies (A. pinsapo) and Pinus (Pinus canariensis). Helical thickenings present in Cathaya, Pseudotsuga (P. sinensis and P. menziesii), some species of Larix (L. decidua, L. griffithii, L. occidentalis, L. potaninii), and Picea (P. abies, P. acoquiana, P. brachytyla, P. breweriana, P. glauca, P. glehnii, P. jezoensis, P. koraiensis, P. mariana, P. maximowiczii, P. morrisonicola, P. purpurea, P. schrenkiana, P. sitchensis, P. smithiana, P. spinulosa and P. torano). Callitroid thickenings absent. Axial parenchyma— present in the whole family, but very scarce, mostly restricted to single cells in marginal arrangement in the last rows of latewood. More abundant in Abietoideae genera (Abies, Cedrus, Keteleeria, Nothotsuga, Pseudolarix and Tsuga). Transverse end walls irregular to nodular. Rays—presence of ray tracheids in all genera of the family except Keteleeria. Always present in Cathaya, Larix, Picea, Pinus, Pseudotsuga and Tsuga, frequent in Cedrus and rare in Abies, Nothotsuga and Pseudolarix. In Pinus, ray tracheid walls are smooth, dentate or reticulate. Ray tracheids with helical thickenings in Cathaya and some species of Larix, Picea and Pseudotsuga. Horizontal walls of ray parenchyma cells pitted, although many species of Pinus do not conform. The same occurs with the end walls of ray parenchyma cells, which are nodular in all genera except in some species of Pinus, and slightly nodular in Pinus section Strobus (e.g., P. cembra, P. koraiensis, P. lambertiana, P. monticola and P. strobus). At family level, cross-field pitting does not follow a common pattern—Abies (piceoid, taxodioid), Cathaya (piceoid), Cedrus (piceoid and taxodioid), Keteleeria (cupressoid and taxodioid), Larix (piceoid), Nothotsuga (cupressoid and taxodioid), Picea (piceoid), Pinus (pinoid and window-like), Pseudolarix (piceoid and taxodioid), Pseudotsuga (piceoid) and Tsuga (piceoid)—but at genus level it can be used to establish groups, e.g., in Pinus. Resin canals— presence of axial and radial resin canals in all non-Abietoideae genera. In Cathaya, Larix, Picea and Pseudotsuga, epithelial cells are thick-walled, and in Pinus, they are thin-walled. In Abietoideae genera, Keteleeria and Nothotsuga have only axial resin canals with thick-walled epithelial cells and Abies, Cedrus, Pseudolarix and Tsuga lack normal resin canals. However, all genera in Pinaceae form traumatic resin canals, both axial and radial in non-Abietoideae genera, and in Abietoideae only traumatic axial resin canals are formed, except in Cedrus, which lacks normal axial and radial resin canals but forms traumatic axial and radial canals. The arrangement of traumatic resin canals is normally in tangential rows, except in Keteleeria and Nothotsuga, where they can also be in solitary arrangement. Mineral inclusions—crystals are frequent in marginal and submarginal cells of ray parenchyma in all genera of Abietoideae and occasional in Pinus and Picea.
Podocarpaceae Endl.— nineteen genera: Acmopyle Pilg. (2 species), Afrocarpus (Buchholz & N.E.Gray) C.N.Page (5), Dacrycarpus (Endl.) de Laub. (9), Dacrydium Sol. ex G.Forst. (22), Falcatifolium de Laub. (6), Halocarpus C.J.Quinn (3), Lagarostrobos Quinn (1), Lepidothamnus Phil. (3), Manoao Molloy (1), Microcachrys Hook.f. (1) Nageia Gaertn. (5), Parasitaxus de Laub. (1), Pherosphaera W.Archer bis (2), Phyllocladus Rich. ex Mirb. (4), Podocarpus L’Hér. ex Pers. (115), Prumnopitys Phil. (3), Retrophyllum C.N.Page (6) Saxegothaea Lindl. (1), Sundacarpus (J.Buchholz & N.E.Gray) C.N.Page (1). Axial tracheids— tracheid pitting in radial walls normally uniseriate. When it is biseriate, arrangement is opposite. Torus defined. Extensions do not occur regularly, but were observed in Lagarostrobos. Notched borders were not observed. The warty layer was not visible. Organic deposits were absent. Helical and callitroid thickenings were absent. Axial parenchyma— present in most genera, in diffuse arrangement, transverse end walls smooth. Absent in Halocarpus, Lagarostrobos, Lepidothamnus, Manoao, Microcachrys and Sundacarpus. Rays— ray tracheids were absent. Horizontal and end walls of ray parenchyma cells were smooth. Cross-field pitting is normally cupressoid and taxodioid, although the genera Lagarostrobos, Manoao, Microstrobos, Microcachrys, Phyllocladus, Prumnopitys (P. andina) and Sundacarpus have window-like pitting in addition to taxodioid. Resin canals— absent. Mineral inclusions—not observed.
Sciadopityaceae Luerss— one genus: Sciadopitys Siebold and Zucc. (1 species) Axial tracheids—tracheid pitting in radial walls uniseriate. Torus was defined. Torus extensions and notched borders were absent. The warty layer was difficult to observe, even with electron microscopy. The deposits were absent; helical and callitroid thickenings were absent. Axial parenchyma—very scarce, virtually absent and transverse end walls were smooth. Rays—ray tracheids were absent. Horizontal and end walls of ray parenchyma cells were smooth. Cross-field pitting window-like. Resin canals—absent. Mineral inclusions—not observed.
Taxaceae Gray—six genera: Amentotaxus Pilg. (6), Austrotaxus Compton (1), Cephalotaxus Siebold and Zucc. ex Endl. (8), Pseudotaxus W.C.Cheng (1), Taxus L. (12), Torreya Arn. (6). Axial tracheids— tracheid pitting in radial walls uniseriate. The torus was defined. Torus extensions and notched borders were absent. The warty layer was not visible. Organic deposits absent. All genera except Austrotaxus have helical thickenings. In Pseudotaxus and Taxus, they are normally single; in Amentotaxus, Cephalotaxus and Torreya, they are single or double; in Torreya, they can be triple. Callitroid thickenings were absent. Axial parenchyma—present regularly in diffuse arrangement in Cephalotaxus, Amentotaxus and Torreya. They were present occasionally in Austrotaxus and rare or absent in Taxus and Pseudotaxus. The transverse end walls were smooth; in Cephalotaxus smooth to irregularly thickened. Rays—ray tracheids were absent. Horizontal and end walls of ray parenchyma cells were smooth. Cross-field pitting cupressoid. Resin canals—absent. Mineral inclusions— not observed.
7. Conclusions
Axial tracheids—The presence of intercellular spaces is frequent in Agathis, Cupressus, Juniperus and Pilgerodendron. The arrangement of pitting is normally uniseriate and when it is biseriate it is generally opposite, except in the three genera of Araucariaceae (Araucaria, Agathis and Wollemia), where pits are closely packed, alternate and hexagonal in appearance. Triseriate arrangement is common in Keteleeria, Sequoia sempervirens, Taiwania cryptomerioides and Taxodium distichum. All conifer wood tracheid pits have a defined torus except for some species of Thuja and Thujopsis. Only Cedrus regularly has scalloped tori. Some decaying wood can show apparently scalloped tori. The presence of torus extensions is frequent in Abies, Actinostrobus, Fitzroya, Juniperus, Lagarostrobos, Pilgerodendron, Tsuga, Thujopsis and Widdringtonia. Notched borders in pits allow for the differentiation of Sequoia (with notched borders) and Sequoiadendron (without). A warty layer is visible under light microscopy in some genera (e.g., Abies, Actinostrobus, Juniperus, Callitris and Tetraclinis) and can be used as a distinguishing feature. Organic deposits can also be used for diagnosis, and are habitual in Araucaria and Agathis. Helical thickenings in axial tracheids are present in all genera of Taxaceae (except Austrotaxus), in Cathaya, Pseudotsuga and in some species of Picea and Larix. Callitroid thickenings, although reported in other genera, can be used for diagnosis in Actinostrobus and Callitris.
Axial parenchyma—the presence of axial parenchyma permits the exclusion of the families Araucariaceae and Sciadopityaceae and the genera Taxus and Pseudotaxus in Taxaceae, Neocallitropsis, Thuja and Xanthocyparis in Cupressaceae, and Halocarpus, Lagarostrobos, Lepidothamnus, Manoao, Microcachrys, Phyllocladus and Sundacarpus in Podocarpaceae. With regard to arrangement, diffuse, tangentially zonate and marginal, even simultaneously, are characteristic of Cupressaceae, while in Podocarpaceae it is diffuse.
Rays—ray composition has high diagnostic value. The presence of ray tracheids is exclusive to the genera of Pinaceae that possess normal resin canals, with the exception, outside this group, of Tsuga and Callitropsis nootkatensis. Tsuga always has ray tracheids, in Cedrus they are frequent, in Abies, Nothotsuga and Pseudolarix they are rare, and in Keteleeria they are absent. The absence of ray tracheids in Keteleeria allows it to be differentiated from Nothotsuga. Other non-Pinaceae species have ray tracheids, particularly in Cupressaceae, e.g., Cupressus arizonica, Sequoia and Thujopsis. The degree of dentation also allows differentiation of groups in the genus Pinus. Helical thickenings in ray tracheids distinguish Cathaya argyrophylla, Pseudotsuga sinensis, Larix potaninii var. himalaica, Picea polita, P. spinulosa and P. torano from the rest of the conifers. Distinctly pitted horizontal walls of radial cells distinguish all the genera of Pinaceae from the other conifer genera, although some species of Pinus do not conform to this, and in Pseudolarix they are less distinct. Nodular end walls are limited to the Pinaceae (absent in some species of Pinus) and Cupressaceae. Cross-field pitting has high diagnostic value, although in some genera more than one type can occur simultaneously, particularly in Cupressaceae.
Resin canals—The presence of normal axial and radial resin canals with thick-walled epithelial cells indicates the group Cathaya, Larix, Picea and Pseudotsuga (Cathaya is distinct from the others because its axial and radial tracheids have helical thickenings); canals with thin-walled epithelial cells are indicative of Pinus. The presence of normal axial resin canals but no radial canals is exclusive to Keteleeria and Nothotsuga. Traumatic axial resin canals are characteristic of Pinaceae and occasionally occur in some Cupressaceae genera (e.g., Microbiota, Sequoia, Sequoiadendron and Tetraclinis). Cathaya, Larix, Picea, Pinus and Pseudotsuga form traumatic axial and radial resin canals. Keteleeria and Nothotsuga form only traumatic axial canals. Abies, Cedrus, Nothotsuga, Pseudolarix and Tsuga lack normal axial and radial resin canals, but all of them can form traumatic axial canals, and Cedrus also forms traumatic radial canals.
Mineral inclusions—the presence of mineral inclusions in Pinaceae, particularly in Abietoideae, has diagnostic value.
Author Contributions
Conceptualization, L.G.E., P.d.P., I.H. and P.G.; Methodology, L.G.E., P.d.P., I.H. and P.G.; Software—Validation, L.G.E. and P.d.P.; Formal Analysis—Investigation, L.G.E., P.d.P., I.H., P.G., A.G.-I. and F.G.-F.; Resources, L.G.E., I.H. and P.G.; Data Curation, -; Writing—Original Draft Preparation, L.G.E., P.d.P., I.H., P.G., A.G.-I. and F.G.-F.; Writing—Review and Editing, L.G.E., P.d.P., I.H. and P.G.; Visualization, A.G.-I. and F.G.-F.; Supervision, L.G.E.; Project Administration, L.G.E., P.d.P., I.H. and P.G.; Funding Acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors are grateful for the contribution of samples from other wood collections that made this study possible.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cole, T.C.H.; Bachelier, J.B.; Hilger, H.H. Tracheophyte Phylogeny Poster—Vascular Plants: Systematics and Characteristics. PeerJ. Prepr. 2019, 7, e2614v3. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2021. Available online: http://www.plantsoftheworldonline.org/ (accessed on 8 December 2021).
- Wang, L.; Wang, D.; Lin, M.M.; Lu, Y.; Jang, X.X.; Jin, B. An embryological study and systematic significance of the primitive gymnosperm Ginkgo biloba. J. Syst. Evol. 2011, 49, 353–361. [Google Scholar] [CrossRef]
- Torelli, N. Dvokrpi ginko (Ginkgo biloba L.) in njegov les [Maidenhair tree (Ginkgo biloba L.) and its wood]. Les 1999, 51, 397–402. [Google Scholar]
- IAWA Committee. IAWA list of microscopic features for softwood identification. IAWA J. 2004, 25, 1–70. [Google Scholar] [CrossRef]
- Terrazas, T. Origin and activity of successive cambia in Cycas (Cycadales). Am. J. Bot. 1991, 78, 1335–1344. [Google Scholar] [CrossRef]
- Carlquist, S. Wood, Bark, and Pith Anatomy of Old World Species of Ephedra and Summary for the Genus. Aliso 1992, 13, 255–295. [Google Scholar] [CrossRef]
- Thompson, W.P. The anatomy and relationships of the Gnetales. I. The genus Ephedra. Ann. Bot. 1912, 26, 1077–1104. [Google Scholar] [CrossRef]
- Bailey, I.W. The development of vessels in angiosperms and its significance in morphological research. Am. J. Bot. 1944, 31, 421–428. [Google Scholar] [CrossRef]
- Muhammad, A.F.; Sattler, R. Vessel structure of Gnetum and origin of angiosperms. Am. J. Bot. 1982, 69, 1004–1021. [Google Scholar] [CrossRef]
- Carlquist, S.; Gowans, D.A. Secondary growth and wood histology of Welwitschia. Bot. J. Linn. Soc. 1995, 118, 107–121. [Google Scholar] [CrossRef]
- Carlquist, S. Wood Anatomy of Gnetales in a Functional, Ecological, and Evolutionary Context. Aliso 2012, 30, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Carlquist, S. Wood, bark, and stem anatomy of gnetales: A summary. Int. J. Plant Sci. 1996, 157, S58–S76. [Google Scholar] [CrossRef]
- Esteban, L.G.; de Palacios, P. Comparative wood anatomy in Abietoideae (Pinaceae). Bot. J. Linn. Soc. 2009, 160, 184–196. [Google Scholar] [CrossRef]
- Gernandt, D.S.; Magallón, S.; López, G.G.; Flores, O.Z.; Willyard, A.; Liston, A. Use of simultaneous analyses to guide fossil-based calibrations of Pinaceae phylogeny. Int. J. Plant Sci. 2008, 169, 1086–1099. [Google Scholar] [CrossRef]
- Farjon, A.; Hiep, N.T.; Harder, D.K.; Loc, P.K.; Averyanov, L. A new genus and species in Cupressaceae (Coniferales) from northern Vietnam, Xanthocyparis vietnamensis. Novon 2002, 12, 179–189. [Google Scholar] [CrossRef]
- Neale, D.B.; Wheeler, N.C. The Conifers: Genomes, Variation and Evolution; Springer: Cham, Switzerland, 2019; p. 590. [Google Scholar] [CrossRef]
- Farjon, A.; Filer, D. An Atlas of the World’s Conifers. An Analysis of Their Distribution, Biogeography, Diversity and Conservation Status; Ed. Brill.: London, UK, 2013; p. 512. [Google Scholar]
- Farjon, A. World Checklist and Bibliography of Conifers, 2nd ed.; Royal Botanic Gardens at Kew: London, UK, 2001. [Google Scholar]
- Sanio, C. Einige Bemerkungen uber den Bau des Holzes. Bot. Z 1860, 18, 193–198, 201–204, 209–217. [Google Scholar]
- Sanio, C. Anatomie der gemeinen Kiefer (Pinus silvestris L.). Jahrb. Wiss. Bot. 1873, 9, 50–126. [Google Scholar]
- Castellarnau, J.M. Estudio Micrográfico del Tallo del Pinsapo (Abies pinsapo Boiss.); Anales de la Sociedad Española de Historia Natural: Madrid, Spain, 1880. [Google Scholar]
- Kleeberg, A. Die Markstrahlen der Coniferen. Bot. Zeit. 1885, 43, 673–686. [Google Scholar]
- Penhallow, D.P. The generic characters of the North American Taxaceae and Coniferae. Mémoires et comptes rendus de la Société Royale du Canada. Proc. Trans. R. Soc. Can. 1896, 2, 33–57. [Google Scholar]
- Jeffrey, E.C. The comparative anatomy and phylogeny of the Coniferales. Part 1. The genus Sequoia. Mem. Boston Soc. Nat. Hist. 1903, 5, 441–459. [Google Scholar]
- Bitting, K.G. The histological difference between Pinus taeda and P. palustris. Proc. Ind. Acad. Sci. 1908, 1, 127–132. [Google Scholar]
- Bailey, I.W. The Structure of the Wood in the Pineae. Bot. Gaz. 1909, 48, 47–55. [Google Scholar] [CrossRef]
- Brooks, F.T.; Stiles, W. The Structure of Podocarpus spinulosus, (Smith) R. Br. Ann. Bot.-Lond. 1910, 24, 305–318. [Google Scholar] [CrossRef]
- Baker, R.T.; Smith, H.G. A Research on the Pines of Australia; The Government of the State of New South Wales, William Applegate Gullick, Government Printer: Sydney, Australia, 1910; p. 458. [Google Scholar]
- Thompson, W.P. The origin of the ray tracheids in the Coniferae. Bot. Gaz. 1910, 50, 101–116. [Google Scholar] [CrossRef]
- Thompson, W.P. Ray tracheids in Abies. Bot. Gaz. 1912, 53, 331–338. [Google Scholar] [CrossRef]
- Gordon, M. Ray Tracheids in Sequoia sempervirens. New Phytol. 1912, 11, 1–6. [Google Scholar] [CrossRef]
- Holden, R. Ray tracheids in the Coniferales. Bot. Gaz. 1913, 55, 56–65. [Google Scholar] [CrossRef]
- Chrysler, M.A. The medullary ray of Cedrus. Bot. Gaz. 1915, 59, 387–396. [Google Scholar] [CrossRef]
- Jeffrey, E.C. The Anatomy of Woody Plants; University of Chicago Press: Chicago, CA, USA, 1917. [Google Scholar]
- Kanehira, R. Anatomical Characters and Identification of Formosan Woods with Critical Remarks from the Climatic Point of View; Bureau of Productive Industries, Government of Formosa: Taipei, Taiwan, 1921. [Google Scholar]
- Kanehira, R. Anatomical Characters and Identification of the Important Woods of the Japanese Empire; Bureau of Productive Industries, Government of Formosa: Taipei, Taiwan, 1926. [Google Scholar]
- Saint-Laurent, J. Études sur les caractères anatomiques des bois d’Algérie. I Bull. Sta. Rech For. N. Afr. 1926, 1, 241–255. [Google Scholar]
- Patton, R.T. Anatomy of Australian coniferous timbers. Proc. R. Soc. Vic. 1927, 40, 2–16. [Google Scholar]
- Pool, D.J.W. On the Anatomy of Araucarian Wood. Recl. Des. Trav. Bot. Neerl. 1929, 25, 482–620. [Google Scholar]
- Metcalfe, C.R. The wood structure of Fokienia hodginsii and certain related coniferae. Bull. Misc. Inf. (R. Bot. Gard. Kew) 1931, 8, 420–425. [Google Scholar] [CrossRef]
- Brown, H.P.; Panshin, A.J. Commercial Timbers of the United States: Their Structure, Identification, Properties, and Uses; McGraw-Hill: New York, NY, USA, 1934. [Google Scholar]
- Brem, M. Anatomical method for determining the wood of the spruce and larch. Bull. Intern. Acad. Sei. Lett. Ser B (I) 1934, 8, 103–111. [Google Scholar]
- Bannan, M.W. Vertical resin ducts in the secondary wood of the Abietineae. New Phytol. 1936, 35, 11–46. [Google Scholar] [CrossRef]
- Peirce, A.S. Anatomy of the Xylem of Pseudolarix. Bot. Gaz. 1934, 95, 667–677. [Google Scholar] [CrossRef]
- Peirce, A.S. Anatomy of the xylem of Sciadopitys. Am. J. Bot. 1935, 22, 895–902. [Google Scholar] [CrossRef]
- Peirce, A.S. Anatomical interrelationships of the Taxodiaceae. Trop. Woods 1936, 46, 1–15. [Google Scholar]
- Peirce, A.S. Systematic anatomy of the woods of the Cupressaceae. Trop. Woods 1937, 49, 5–21. [Google Scholar]
- Shimakura, M. Anatomy of the wood of Taiwania. Bot. Mag. 1937, 51, 694–700. [Google Scholar] [CrossRef]
- Bernath, E.L. Coniferous forest trees of Chile. Trop. Woods 1937, 52, 19–26. [Google Scholar]
- Covas, G. Las coníferas indígenas de la República Argentina. Rev. Fac. Agron. B Aires 1938, 21, 201–203. [Google Scholar]
- Phillips, E.W.J. The identification of coniferous wood by their microscopic structure. Bot. J. Linn. Soc. 1941, 52, 259–320. [Google Scholar] [CrossRef]
- Greguss, P. Identification of Living Gymnosperms on the Basis of Xylotomy; Akadémiai Kiado: Budapest, Hungary, 1955; p. 263. [Google Scholar]
- Kukachka, F. Identification of coniferous wood. Tappi 1960, 43, 887–896. [Google Scholar]
- Barefoot, A.; Hankins, F.W. Identification of Modern and Tertiary Woods; Oxford University Press: New York, NY, USA, 1982; p. 189. [Google Scholar]
- Esteban, L.G.; de Palacios, P.; Guindeo, A.; García, L.; Lázaro, I.; González, L.; Rodríguez, Y.; García, F.; Bobadilla, I.; Camacho, A. Anatomy and Identification of Conifers Wood as a Species; Fundación Conde del Valle de Salazar—Mundi Prensa: Madrid, Spain, 2002; p. 421. [Google Scholar]
- Heinz, I. Systematische Erfassung und Dokumentation der mikroanatomischen Merkmale der Nadelhölzer aus der Klasse der Pinatae. Ph.D. Thesis, Technische Universität München, Munich, Germany, 2004; p. 209.
- Clarke, S.H. A multiple-entry perforated-card key with special reference to the identification of hardwoods. New Phytol. 1938, 37, 369–374. [Google Scholar] [CrossRef]
- Kennedy, R.W.; Sastry, C.B.R.; Barton, G.M.; Ellis, E.L. Crystals in wood of genus Abies indigenous to Canada and United States. Can. J. Bot. 1968, 46, 1221–1228. [Google Scholar] [CrossRef]
- Sudo, S. Anatomical studies on the wood of species of Picea, with some considerations on their geographical distribution and taxonomy. Bull. Govt. Exp. Stat. 1968, 215, 39–130. [Google Scholar]
- Werker, E.; Fahn, A. Resin ducts of Pinus halepensis MilI.—Their structure, development and pattern of arrangement. Bot. J. Linn. Soc. 1969, 62, 379–411. [Google Scholar] [CrossRef]
- Bauch, J.; Liese, W.; Schultze, R. The morphological variability of the bordered pit membranes in gymnosperms. Wood Sci. Technol. 1972, 6, 165–184. [Google Scholar] [CrossRef]
- Bosshard, H.H. Holzkunde—Mikroskopie und Makroskopie des Holzes, Band 1; Birkhauser, Basel: Stuttgart, Germany, 1974. [Google Scholar]
- Core, H.A.; Cóté, W.A.; Day, A.C. Wood structure and identification; Syracuse University Press: New York, NY, USA, 1979. [Google Scholar]
- Chavchavadze, E.S. Wood of Conifers (Drevesina khvoinykh); Original in Russian; Akad. Nauk SSSR: Moscow-Leningrad, Russia, 1979. [Google Scholar]
- Bartholin, T. The Picea-Larix problem. IAWA Bull. 1979, 1, 7–10. [Google Scholar]
- Panshin, A.J.; De Zeeuw, C. Textbook of Wood Technology, 4th ed.; McGraw-Hill: New York, NY, USA, 1980. [Google Scholar]
- Baas, P.; Schmid, R.; van Heuven, B.J. Wood anatomy of Pinus longaeva (bristlecone pine) and the sustained Iength-on-age increase of its tracheids. IAWA Bull. 1986, 7, 221–228. [Google Scholar] [CrossRef]
- Suzuki, M.; Noshiro, S. Wood structure of Himalayan plants. In Himalayan Plants; Ohba, H., Malla, S.B., Eds.; The University Museum, the University of Tokyo, Bulletin Tokyo: Tokyo, Japan, 1988; Volume 31, pp. 341–379. [Google Scholar]
- LaPasha, C.A.; Wheeler, E.A. Resin canals in Pinus taeda: Longitudinal canal lengths and interconnections between longitudinal and radial canals. IAWA Bull. 1990, 11, 227–238. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Anatomy of European Woods; Paul Haupt: Berne, Switzerland, 1990. [Google Scholar]
- Roig, F.A. Comparative wood anatomy of southern South American Cupressaceae. IAWA Bull. 1992, 13, 151–162. [Google Scholar] [CrossRef]
- Vogel, K. Mikroskopische Untersuchung zur Typisierung der Kreuzungsfeldtupfel bei Nadelholzern. Diplomarbeit der Forstwissenschaftlichen. Ph.D. Thesis, Fakultat der Ludwig-Maximilians- Universitat München, Munich, Germany, 1995. [Google Scholar]
- Ilic, J. Separation of the woods of Callitris glaucophylla (white cypress pine) and C. endlicheri (black cypress pine). Recent Advances in Wood Anatomy. In Proceedings of the IAWA Wood Anatomy Conference, Rotorua, New Zealand, 20–24 November 1994. [Google Scholar]
- Kondo, Y.; Fujii, T.; Hayashi, Y.; Kato, A. Organic crystals in the tracheids of Torreya yunnanensis. IAWA J. 1996, 17, 393–403. [Google Scholar] [CrossRef]
- Heinz, I. Entwicklung von Systemkomponenten für die computerunterstützte Bestimmung von Nadelhölzern in DELTA/INTKEY. Diplomarbeit. Ph.D. Thesis, Universitat Hamburg, Hamburg, Germany, 1997. [Google Scholar]
- Kellogg, R.M.; Rowe, S.; Koeppen, R.C.; Miller, R.B. Identification of the wood of the soft pines of western North America. IAWA Bull. 1982, 3, 95–101. [Google Scholar] [CrossRef]
- Yoshizawa, N.; Itoh, T.; Shimaji, K. Helical thickenings in normal and compression wood of some softwoods. IAWA Bull. 1985, 6, 131–138. [Google Scholar] [CrossRef]
- Esteban, L.G.; Guindeo, A.; de Palacios, P. Maderas de Coníferas. Anatomía de Géneros; Fundación Conde del Valle de Salazar: Madrid, Spain, 1996; p. 336. [Google Scholar]
- Heady, R.D.; Evans, P.D. Callitroid (callitrisoid) thickening in Callitris. IAWA J. 2000, 21, 293–319. [Google Scholar] [CrossRef]
- Heady, R.D.; Banks, J.D.; Evans, P.D. Wood anatomy of Wollemi Pine (Wollemia nobilis, Araucariaceae). IAWA J. 2002, 23, 339–358. [Google Scholar] [CrossRef]
- Wiedenhoeft, A.C.; Miller, R.B. Brief comments on the nomenclature of softwood axial resin canals and their associated cells. IAWA J. 2002, 23, 299–303. [Google Scholar] [CrossRef]
- Visscher, G.E.; Jagels, R. Separation of Metasequoia and Glyptostrobus (Cupressaceae) based on wood anatomy. IAWA J. 2003, 24, 439–451. [Google Scholar] [CrossRef]
- Xiaomei, J. Atlas of Gymnosperms Woods of China; Science Press Ltd.: Beijing, China, 2010; p. 490. [Google Scholar]
- de Palacios, P.; Esteban, L.G.; García Fernández, F.; García-Iruela, A.; Conde, M.; Román-Jordán, E. Comparative wood anatomy of Juniperus from Macaronesia. IAWA J. 2014, 35, 186–198. [Google Scholar] [CrossRef]
- Román-Jordán, E. Anatomía comparada de la madera de Cupressaceae y su correspondencia con los estudios de filogenia. Ph.D. Thesis, Universidad Politécnica de Madrid, Madrid, Spain, 4 April 2016. [Google Scholar]
- Troncoso, O.; Greslebin, A. Trabeculae in Patagonian mountain cypress (Austrocedrus chilensis) associated with Phytophthora austrocedri infection. IAWA J. 2018, 39, 209–220. [Google Scholar] [CrossRef]
- Purusatama, B.D.; Kim, N.H. Cross-field pitting characteristics of compression, lateral, and opposite wood in the stem wood of Ginkgo biloba and Pinus densiflora. IAWA J. 2020, 41, 48–60. [Google Scholar] [CrossRef]
- InsideWood. 2004-Onwards. Available online: http://insidewood.lib.ncsu.edu/search (accessed on 11 October 2021).
- Watson, L.; Dallwitz, M.J. 2008 Onwards. The Families of Gymnosperms. Version: 5 August 2019. Available online: delta-intkey.com (accessed on 11 October 2021).
- Earle, C.J. The Gymnosperm Database, 2001 Owards. Available online: http://www.conifers.org (accessed on 20 November 2022).
- Galtier, J.; Meyer-Berthaud, B. The diversification of early arborescent seed ferns. J. Torrey Bot. Soc. 2006, 133, 7–19. [Google Scholar] [CrossRef]
- Brown, H.P.; Panshin, A.J.; Forsaith, C.C. Textbook of Wood Technology. Vol. I. Structure, Identification, Defects and Uses of the Commercial Woods of the United States; McGraw-Hill Book Company: New York, NY, USA, 1949; 652p. [Google Scholar]
- Esteban, L.G.; Guindeo, A.; Peraza, C.; de Palacios, P. La Madera y su Anatomía; Fundación Conde del Valle de Salazar, Ed.; Mundi-Prensa y AiTiM: Madrid, Spain, 2003; 327p. [Google Scholar]
- Wilson, J.P.; Knoll, A.H. A physiologically explicit morphospace for tracheid-based water transport in modern and extinct seed plants. Paleobiology 2010, 36, 335–355. [Google Scholar] [CrossRef]
- Decombeix, A.-L.; Boura, A.; Tomescu, A.M.F. Plant hydraulic architecture through time: Lessons and questions on the evolution of vascular systems. IAWA J. 2019, 40, 387–420. [Google Scholar] [CrossRef]
- Ladell, J.T. A new method of measuring tracheid length. Forestry 1959, 32, 124–125. [Google Scholar] [CrossRef]
- Wilkins, A.P.; Bamber, R.K. A comparison between Ladell’s wood section method and the macerated wood method for tracheid length determination. IAWA Bull. 1983, 4, 245–247. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Dichteschwankungen in Jahrringen von Nadelhölzern in Beziehung zu klimatisch-ökologischen Faktoren, oder das Problem der Falschen Jahrringe, Bericht Nr. 213; Eidgenössische Anstalt für das forstliche Versuchswesen: Brimensdorf, Switzerland, 1980. [Google Scholar]
- Schweingruber, F.H. Tree Rings and Environment. Dendroecology; Paul Haupt: Berne, Switzerland, 1996; pp. 71–93. [Google Scholar]
- Willebrand, G. Untersuchung von ausgewählten mikroanatomischen Merkmalen zur Bestimmung von Nadelhölzern. Diplomarbeit. Ph.D. Thesis, Fachhochschule Rosenheim, Fachbereich Holztechnik, Rosenheim, Germany, 1995. [Google Scholar]
- Mio, S.; Matsumoto, T. Morphological observation on longitudinal intercellular spaces in normal softwoods. Bull. Kyushu Univ. For. 1979, 51, 13–18, (In Japanese with English abstract). [Google Scholar]
- Nagai, S.; Utsumi, Y. The function of intercellular spaces along the ray parenchyma in sapwood, intermediate wood, and heartwood of Cryptomeria japonica (Cupressaceae). Am. J. Bot. 2012, 99, 1553–1561. [Google Scholar] [CrossRef]
- Beck, C.B.; Wight, D.C. Progymnosperms. In Origin and Evolution of Gymnosperms; Beck, C.B., Ed.; Columbia University Press: New York, NY, USA, 1988; pp. 1–84. [Google Scholar]
- Prestianni, C.; Decombeix, A.-L.; Thorez, J.; Fokand, D.; Gerrienne, P. Famennian charcoal of Belgium. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 291, 60–71. [Google Scholar] [CrossRef]
- Decombeix, A.-L.; Meyer-Berthaud, B.; Galtier, J.; Talent, J.; Mawson, R. Diversity of arborescent lignophytes in the Tournaisian vegetation of Queensland (Australia): Paleoecological and paleogeographical significance. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 301, 39–55. [Google Scholar] [CrossRef]
- Morrow, A.C.; Dute, R.R. Development and structure of pit membranes in the rhizome of the woody fern Botrychium dissectum. IAWA J. 1998, 19, 429–441. [Google Scholar] [CrossRef]
- Bodnar, J.; Ruiz, D.P.; Artabe, A.E.E.; Morel, E.M.; Ganuza, D.G. Voltziales y Pinales (= Coniferales) de la Formación Cortaderita (Triásico Medio), Argentina, y su implicancia en la reconstrucción de las coníferas triásicas. Rev. Bras. Paleontol. 2015, 18, 141–160. [Google Scholar] [CrossRef]
- Jansen, S.; Lamy, J.B.; Burlett, R.; Cochard, H.; Gasson, P.; Delzon, S. Plasmodesmatal pores in the torus of bordered pit membranes affect cavitation resistance of conifer xylem. Plant Cell Env. 2012, 35, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Harada, H.; Miyazaki, Y.; Wakashima, T. Electron microscopic investigation on the cell wall structure of wood. Bull. Govt. For. Exp. Stn. 1968, 104, 1–115. [Google Scholar]
- Liese, W. Elektronenmikroskopie des Holzes. In Handbuch der Mikroskopie in der Technik; Freund, H., Ed.; Umschau-Verlag: Frankfurt, Germany, 1970; pp. 109–170. [Google Scholar]
- Sano, Y.; Kawakami, Y.; Ohtani, J. Variation in the structure of intertracheary pit membranes in Abies sachalinensis, as observed by field-emission electron microscopy. IAWA J. 1999, 20, 375–388. [Google Scholar] [CrossRef]
- Esteban, L.G.; de Palacios, P.; García Fernández, F.; Moreno, R. Wood anatomy of the genus Abies: A review. IAWA J. 2009, 30, 231–245. [Google Scholar] [CrossRef]
- Liese, W. The warty layer. In Cellular Infrastructure of Woody Plants; Côté, W.A., Ed.; Syracuse University Press: Syracuse, NY, USA, 1965; pp. 251–269. [Google Scholar]
- Heady, R.D.; Evans, P.D. Wood anatomy of Actinostrobus (Cupressaceae). IAWA J. 2005, 26, 79–92. [Google Scholar] [CrossRef]
- Esteban, L.G.; de Palacios, P.; García-Iruela, A.; Román-Jordán, E.; García-Fernández, F.; Díaz Fernández, S.; Conde, M. Wood anatomy of Tetraclinis articulata from its natural distribution area in southeast Spain. IAWA J. 2015, 36, 22–35. [Google Scholar] [CrossRef]
- Jansen, S.; Smets, E.; Baas, P. Vestures in woody plants: A review. IAWA J. 1998, 19, 347–382. [Google Scholar] [CrossRef]
- Zimmerman, M.H. Xylem Structure and the Ascent of Sap; Springer: New York, NY, USA, 1983. [Google Scholar]
- Peraza, C. Estudio de las Maderas de Coníferas Españolas y de la Zona Norte de Marruecos; IFIE: Madrid, Spain, 1964. [Google Scholar]
- Esteban, L.G.; Gasson, P.; Climent, J.M.; de Palacios, P.; Guindeo, A. The wood of Pinus canariensis and its resinous heartwood. IAWA J. 2005, 26, 69–77. [Google Scholar] [CrossRef]
- Hillis, W.E. Heartwood and Tree Exudates; Springer: Berlin, Germany, 1987. [Google Scholar] [CrossRef]
- Funada, R.; Miura, H.; Shibagaki, M.; Furusawa, O.; Miura, T.; Fukatsu, E.; Kitin, P. Involvement of Localized Cortical Microtubules in the Formation of a Modified Structure of Wood. J. Plant Res 2001, 114, 491–497. [Google Scholar] [CrossRef]
- Xie, F.H. Wood Structure and Taxonomical Position of Cathaya (Abstract). Symposium on the Explotation of Tropical Resources in South China; Forestry Press: Beijing, China, 1957. [Google Scholar]
- Yatsenko-Khmelevsky, A.A.; Budkevich, E.V. On the Wood anatomy of Cathaya argyrophylla Chun et Kuang (Pinaceae). Bot. Zurn. 1958, 43, 477–480. [Google Scholar]
- Greguss, P. Similar xylotomy and leaf-epidermis of the Pseudotsuga and the new genus Cathaya. Bot. Kozlem. 1970, 57, 51–55. [Google Scholar]
- Greguss, P. Xylotomy of the Living Conifers; Akadémiai Kiadó: Budapest, Hungary, 1972; p. 329. [Google Scholar]
- Cheng, T.C. Tropical and Subtropical Woods in China (Identifications, Properties and Uses); Beijing Science Press: Beijing, China, 1980. [Google Scholar]
- Hu, Y.S.; Wang, F.H. Anatomical studies of Cathaya (Pinaceae). Am J. Bot. 1984, 71, 727–735. [Google Scholar] [CrossRef]
- Esteban, L.G.; de Palacios, P.; García-Iruela, A.; García-Fernández, F.; García-Esteban, L.; González de Vega, D. Comparative anatomy of wood in Pinaceae with reference to its systematic position. Forests 2021, 12, 1706. [Google Scholar] [CrossRef]
- Anagnost, S.E.; Meyer, R.W.; De Zeeuw, C. Confirmation and significance of Bartholin’s method for identification of the wood of Picea and Larix. IAWA J. 1994, 15, 171–184. [Google Scholar] [CrossRef]
- Yatsenko-Khmelevsky, A.A. The Principles and Methods of Anatomical Investigation of Wood (in Russian); Akad. Naúk SSSR: Moscow-Leningrad, Russia, 1954; p. 337. [Google Scholar]
- Phillips, E.W.J. Identification of Softwoods by their Microscopic Structure; Forest Products Research Bull. No. 22; HMSO Department of Scientific and Industrial Research: London, UK, 1948. [Google Scholar]
- Keunecke, D. Elasto-mechanical characterisation of yew and spruce wood with regard to structure-property relationships. Ph.D. Thesis, Eidgenössische Technische Hochschule ETH Zürich, Zürich, Switzerland, 2008. [Google Scholar] [CrossRef]
- Howard, E.T.; Manwiller, F.G. Anatomical characteristics of southern pine stemwood. Wood Sci. 1969, 2, 77–86. [Google Scholar]
- Meylan, B.A.; Butterfield, B.G. The Structure of New Zealand Woods, DSIR Bull. 222; NZ Department of Scientific and Industrial Research: Wellington, New Zealand, 1978. [Google Scholar]
- Carlquist, S. Wood anatomy of Cynareae (Compositae). Aliso 1965, 6, 13–24. [Google Scholar] [CrossRef]
- McElhanney, T.A. Associates in the Forest Products Laboratories of Canada. Canadian Woods: Their Properties and Uses; Department of the Interior: Ottawa, Canada, 1935; p. 345. [Google Scholar]
- Grosser, D. On the occurrence of trabeculae with special consideration of diseased trees. IAWA Bull. 1986, 7, 319–341. [Google Scholar] [CrossRef]
- Ghimire, B.; Lee, C.; Heo, K. Comparative wood anatomy of Taxaceae. Aust Syst Bot. 2015, 28, 160–172. [Google Scholar] [CrossRef]
- Gasson, P.; Baas, P.; Wheeler, E.A. Wood anatomy of CITES-listed tree species. IAWA J. 2011, 32, 155–198. [Google Scholar] [CrossRef]
- Galtier, J. On the earliest arborescent gymnosperms. Cour. Forsch. Senckenberg 1992, 147, 119–125. [Google Scholar]
- Dunn, M.T. A review of permineralized seed fern stems of the Upper Paleozoic. J. Torrey Bot. Soc. 2006, 133, 20–32. [Google Scholar] [CrossRef]
- Esteban, L.G.; de Palacios, P.; Guindeo, A.; Fernandez, F.G. Comparative anatomy of the wood of Abies pinsapo and its two Moroccan varieties. IAWA J. 2007, 28, 285–299. [Google Scholar] [CrossRef]
- Hudson, R.H. The anatomy of the genus Pinus in relation to its classification. J. Inst. Wood Sci. 1960, 6, 26–46. [Google Scholar]
- Jacquiot, C. Atlas D’anatomie des Bois des Conifères; Institut National du Bois: Paris, France, 1955; p. 197. [Google Scholar]
- Jane, F.W. The structure of Wood, 2nd ed.; A & C Black Publishers Ltd.: London, UK, 1970; p. 478. [Google Scholar]
- Rol, R. Note sur un essai de classification du genre Pinus d’après des caractères tirés de l’anatomie du bois. Rapp Congr Soco Sayo 1932, 65, 333–341. [Google Scholar]
- Kibblewhite, R.P.; Thompson, N.S. The ultrastructure of the middle lamella region in resin canal tissue isolated from slash pine holocellulose. Wood Sci. Technol. 1973, 7, 112–126. [Google Scholar] [CrossRef]
- Esau, K. Anatomy of Seed Plants; Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Fahn, A. Secretory Tissues in Plants; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Ickert-Bond, S.M. Reexamination of wood anatomical features in Pinus krempfii (Pinaceae). IAWA J. 2001, 22, 355–365. [Google Scholar] [CrossRef]
- Takahara, S.; Nobuchi, T.; Harada, H.; Saiki, H. Cell arrangement in the tissue surrounding axial resin canals in the wood of European spruce. Mokuzai Gakkaishi 1982, 28, 197–201. [Google Scholar]
- Sato, K.; Ishida, S. Resin canals in the wood of Larix leptolepis Gord. II. Morphology of vertical resin canals. Res. Bull. Coll. Exp. 1982, 39, 297–326. [Google Scholar]
- Jain, K.K. Evolution of wood structure in Pinaceae. Isr. J. Bot. 1976, 25, 28–33. [Google Scholar]
- Wu, H.; Hu, Z.H. Comparative anatomy of resin ducts of the Pinaceae. Trees 1997, 11, 135–143. [Google Scholar] [CrossRef]
- Lin, J.; Liang, E.; Farjon, A. The occurrence of vertical resin canals in Keteleeria, with reference to its systematic position in Pinaceae. Bot. J. Linn Soc 2000, 134, 567–574. [Google Scholar] [CrossRef]
- Jeffrey, A.H. A cladistic analysis of Conifers: Preliminary results. J. Arnold Arbor. 1987, 68, 269–307. [Google Scholar]
- Lin, J.X.; Hu, Y.S.; Wang, F.H. Wood and bark anatomy of Nothotsuga (Pinaceae). Ann. Mo. Bot. Gard. 1995, 82, 603–609. [Google Scholar]
- Grosser, D. Die Hölzer Mitteleuropas; Springer: New York, NY, USA, 1977. [Google Scholar]
- Schweingruber, F.H. Microscopic Wood Anatomy; Swiss Federal Institute of Forestry Research: Birmensdorf, Switzerland, 1978. [Google Scholar]
- Hoadley, R.B. Wood Identification: Accurate Results with Simple Tools; Taunton Press: Newtown, CT, USA, 1990. [Google Scholar]
- Dickison, W.C. Integrative Plant Anatomy; Academic Press: New York, NY, USA, 2000. [Google Scholar]
- Lotova, L.L. On the Correlation of the Anatomical Features of the Wood and Phloem in the Pinaceae. J. Moscow Univ. 1975, 1, 41–51. [Google Scholar]
- Bailey, I.W.; Faull, A.F. The cambium and its derivative tissues. IX. Structural variability in redwood. Sequoia sempervirens and its significance in the identification of fossil woods. J. Arnold Arbor. 1934, 15, 233–254. [Google Scholar]
- Benkova, V.E.; Schweingruber, F.H. Anatomy of Russian woods. An Atlas for the Identification of Trees, Shrubs, Dwarf Shrubs and Woody Lianas from Russia; Haupt Verlag: Bern, Switzerland, 2004; p. 456. [Google Scholar]
- Anderson, A.P. Comparative anatomy of the normal and diseased organs of Abies balsamea affected with Aecidium elatinum. Bot. Gaz. 1897, 24, 309–344. [Google Scholar] [CrossRef]
- Jeffrey, E.C. The comparative anatomy and phylogeny of the Coniferales. Part. 2. The Abietineae. Mem. Boston Soc. Nat. Hist. 1905, 6, 38–44. [Google Scholar]
- Chamberlain, C.J. Gymnosperms. Structure and Evolution; The University of Chicago Press: Chicago, IL, USA, 1935; p. 484. [Google Scholar]
- Pearson, R.S.; Brown, H.P. Commercial Timbers of India; Government of India Central Publication Branch: Calcutta, India, 1932; Volume 2. [Google Scholar]
- Penhallow, D.P. North American Gymnosperms; Ginn & Co.: Boston, MA, USA, 1907; p. 374. [Google Scholar]
- Vierhapfer, F. Entwurf eines neuen Systemes der Coniferen; Abhandl der KK Zool botan Gesellschaft in Wien: Jena, Germany, 1910; p. 56. [Google Scholar]
- Record, S. Identification of the Economic Woods of the United States; J. Wiley: New York, NY, USA, 1919; p. 157. [Google Scholar]
- Wiedenhoeft, A.C.; Miller, R.B.; Theim, T.J. Analysis of three microscopic characters for separating the wood of Pinus contorta and P. Ponder. IAWA J. 2003, 24, 257–267. [Google Scholar] [CrossRef]
- Krahmer, R.L.; Hemingway, R.W.; Hillis, W.E. The cellular distribution of lignans in Tsuga heterophylla wood. Wood Sci. Technol. 1970, 4, 122–139. [Google Scholar] [CrossRef]
- Díaz-Vaz, J.E. Anatomía de Maderas; Marisa Cuneo Ediciones: Valdivia, Chile, 2003. [Google Scholar]
- Blanco, M.L.; Carpio, I.M.; Muñoz, F. Fichas Técnicas de Veinte Especies Maderables de Importancia Comercial en Costa Rica, 1st ed.; Editorial Universidad de Costa Rica: San Jose de Costa Rica, Costa Rica, 2005. [Google Scholar]
- Yang, K.C.; Yang, Y.H. Minute Structure of Taiwanese Woods; Hua Shiang Yuan Publishing Co.: Taipei, Taiwan, 1987. [Google Scholar]
- Tang, Y. Timber studies of Chinese trees IV. Anatomical studies and identification of Chinese softwoods I. Bull. Fan Mem. Inst. Biol. 1933, 4, 309–368. [Google Scholar]
- Gamble, J.S. A Manual of INDIAN TIMBERS: An Account of the Growth, Distribution, and Uses of the Trees and Shrubs of India and Ceylon with Descriptions of Their Wood-Structure; Sampson Low, Marston & Company: London, UK, 1902. [Google Scholar]
- Kato, K. A trial to detect optimal pin-pricking timing in evaluating the ability to form traumatic resin canals of Cryptomeria japonica for selecting resistant trees to Semanotus japonicus (Coleoptera: Cerambycidae). J. For. Res.-Jpn. 2008, 13, 386–392. [Google Scholar] [CrossRef]
- Fang, W.; Wu, Y. Anatomical Properties and Colorized Illustrations of Important Commercial Wood Species of Hunan in China; Science Press: Beijing, China, 2010. [Google Scholar]
Figure 1.
Gymnosperm tree based on molecular phylogeny data of tracheophytes [1].
Figure 1.
Gymnosperm tree based on molecular phylogeny data of tracheophytes [1].

Figure 2.
Coniferous gymnosperms. (A) Fitzroya cupressoides (Molina) I.M.Johnst.; (B) Mixed conifer forest of Abies pinsapo subsp. marocana (Trab.) Emb. and Maire, Cedrus atlantica (Endl.) Manetti ex Carrière and Pinus nigra subsp. salzmannii (Dunal) Franco; (C) Juniperus cedrus, Webb and Berthel.; (D) Araucaria araucana (Molina) K.Koch; (E) Juniperus canariensis, Guyot and Mathou.
Figure 2.
Coniferous gymnosperms. (A) Fitzroya cupressoides (Molina) I.M.Johnst.; (B) Mixed conifer forest of Abies pinsapo subsp. marocana (Trab.) Emb. and Maire, Cedrus atlantica (Endl.) Manetti ex Carrière and Pinus nigra subsp. salzmannii (Dunal) Franco; (C) Juniperus cedrus, Webb and Berthel.; (D) Araucaria araucana (Molina) K.Koch; (E) Juniperus canariensis, Guyot and Mathou.

Figure 3.
Non-coniferous gymnosperms. (A) Cycas revoluta Thunb.; (B) Ginkgo biloba L.; (C) Zamia sp.
Figure 3.
Non-coniferous gymnosperms. (A) Cycas revoluta Thunb.; (B) Ginkgo biloba L.; (C) Zamia sp.

Figure 4.
Non-coniferous gymnosperms. (A) Ephedra viridis Coville; (B) Welwitschia mirabilis Hook. f.; (C) Gnetum gnemon L.
Figure 4.
Non-coniferous gymnosperms. (A) Ephedra viridis Coville; (B) Welwitschia mirabilis Hook. f.; (C) Gnetum gnemon L.

Figure 5.
Anatomy of the “wood” of non-coniferous gymnosperms. (A–C) Ephedra gerardiana Wall. ex Klotzsch and Garcke; (D–F) Ginkgo biloba L.; (G–I) Gnetum gnemon L.
Figure 5.
Anatomy of the “wood” of non-coniferous gymnosperms. (A–C) Ephedra gerardiana Wall. ex Klotzsch and Garcke; (D–F) Ginkgo biloba L.; (G–I) Gnetum gnemon L.

Figure 6.
Worldwide distribution of conifers (shown in black) (based on Farjon and Filer [18]).
Figure 6.
Worldwide distribution of conifers (shown in black) (based on Farjon and Filer [18]).

Figure 7.
Growth rings. (A) Distinct growth ring boundaries in Pinus cembra L.; (B) Indistinct growth ring boundaries in Afrocarpus gracilior (Pilg.) C.N.Page; (C) Growth rings with abrupt transition in Abies nordmanniana subsp. equi-trojani (Asch. & Sint. ex Boiss.) Coode and Cullen; (D) Growth rings with gradual transition in Abies alba Mill. Scale bars. (A–D) = 500 μm.
Figure 7.
Growth rings. (A) Distinct growth ring boundaries in Pinus cembra L.; (B) Indistinct growth ring boundaries in Afrocarpus gracilior (Pilg.) C.N.Page; (C) Growth rings with abrupt transition in Abies nordmanniana subsp. equi-trojani (Asch. & Sint. ex Boiss.) Coode and Cullen; (D) Growth rings with gradual transition in Abies alba Mill. Scale bars. (A–D) = 500 μm.

Figure 8.
Growth rings. (A) Normal rings in Larix sibirica Ledeb.; (B) Undulating rings in Pilgerodendron uviferum (D.Don) Florin; (C) Hazel growth rings in Picea abies (L.) H.Karst.; (D) False growth ring in Pinus sylvestris L.; (E) Wedging rings in Nageia nagi (Thunb.) Kuntze. Scale bars. (A) = 50 μm; (B,C,E) = 1 mm; (D) = 250 μm.
Figure 8.
Growth rings. (A) Normal rings in Larix sibirica Ledeb.; (B) Undulating rings in Pilgerodendron uviferum (D.Don) Florin; (C) Hazel growth rings in Picea abies (L.) H.Karst.; (D) False growth ring in Pinus sylvestris L.; (E) Wedging rings in Nageia nagi (Thunb.) Kuntze. Scale bars. (A) = 50 μm; (B,C,E) = 1 mm; (D) = 250 μm.

Figure 9.
Axial tracheids. (A) Thin-walled in Pinus strobus L.; (B) Thick-walled in Pinus palustris Mill.; (C) Polygonal tracheids in Callitropsis nootkatensis (D.Don) Oerst. (Syn.—Xanthocyparis nootkatensis (D.Don) Farjon & Harder); (D) Circular tracheids in Callitris endlicheri (Parl.) F.M.Bailey (SEM); (E) Polygonal tracheids with circular lumen in Falcatifolium falciforme (Parl.) de Laub. Scale bars. (A,B) = 250 μm; (C–E) = 50 μm.
Figure 9.
Axial tracheids. (A) Thin-walled in Pinus strobus L.; (B) Thick-walled in Pinus palustris Mill.; (C) Polygonal tracheids in Callitropsis nootkatensis (D.Don) Oerst. (Syn.—Xanthocyparis nootkatensis (D.Don) Farjon & Harder); (D) Circular tracheids in Callitris endlicheri (Parl.) F.M.Bailey (SEM); (E) Polygonal tracheids with circular lumen in Falcatifolium falciforme (Parl.) de Laub. Scale bars. (A,B) = 250 μm; (C–E) = 50 μm.

Figure 10.
Intercellular spaces. (A) Agathis alba (Rumph. ex Hassk.) Foxw.; (B) Pilgerodendron uviferum (D.Don); (C) Intercellular spaces and spiral grooves in compression wood in Taiwania cryptomerioides Hayata (SEM). Scale bars. (A–C) = 50 μm.
Figure 10.
Intercellular spaces. (A) Agathis alba (Rumph. ex Hassk.) Foxw.; (B) Pilgerodendron uviferum (D.Don); (C) Intercellular spaces and spiral grooves in compression wood in Taiwania cryptomerioides Hayata (SEM). Scale bars. (A–C) = 50 μm.

Figure 11.
Tangential pits. (A) Tangential pits in transverse section in Chamaecyparis obtusa (Siebold & Zucc.) Endl. (SEM); (B) Tangential pits in transverse section in Athrotaxis selaginoides D.Don; (C) Tangential pits in tangential section in Tetraclinis articulata (Vahl) Mast. Scale bars. (A–C) = 50 μm.
Figure 11.
Tangential pits. (A) Tangential pits in transverse section in Chamaecyparis obtusa (Siebold & Zucc.) Endl. (SEM); (B) Tangential pits in transverse section in Athrotaxis selaginoides D.Don; (C) Tangential pits in tangential section in Tetraclinis articulata (Vahl) Mast. Scale bars. (A–C) = 50 μm.

Figure 12.
Tracheid pits. (A) Juniperus barbadensis L. (SEM), 1. Aperture; 2. Dome or chamber; 3. Torus; (B) Callitropsis nootkatensis (D.Don) Oerst. (SEM), 1. Dome or chamber; 2. Aperture; (C) Hesperocyparis sargentii (Jeps.) Bartel (Syn.—Cupressus sargentii Jeps.) (SEM). 1. Torus; 2. Margo. Scale bars. (A) = 10 μm; (B,C) = 5 μm.
Figure 12.
Tracheid pits. (A) Juniperus barbadensis L. (SEM), 1. Aperture; 2. Dome or chamber; 3. Torus; (B) Callitropsis nootkatensis (D.Don) Oerst. (SEM), 1. Dome or chamber; 2. Aperture; (C) Hesperocyparis sargentii (Jeps.) Bartel (Syn.—Cupressus sargentii Jeps.) (SEM). 1. Torus; 2. Margo. Scale bars. (A) = 10 μm; (B,C) = 5 μm.

Figure 13.
Pit arrangement. (A) Uniseriate in Abies fraseri (Pursh) Poir.; (B) Biseriate in opposite arrangement in Pinus arizonica Engelm.; (C) Multiseriate in Taxodium distichum (L.) Rich.; (D) Biseriate alternate and polygonal in Araucaria columnaris (G.Forst.) Hook.; (E) Pits in radial wall less than half tracheid diameter in Dacrydium elatum (Roxb.) Wall. ex Hook. Scale bars. (A–D) = 50 μm; (E) = 25 μm.
Figure 13.
Pit arrangement. (A) Uniseriate in Abies fraseri (Pursh) Poir.; (B) Biseriate in opposite arrangement in Pinus arizonica Engelm.; (C) Multiseriate in Taxodium distichum (L.) Rich.; (D) Biseriate alternate and polygonal in Araucaria columnaris (G.Forst.) Hook.; (E) Pits in radial wall less than half tracheid diameter in Dacrydium elatum (Roxb.) Wall. ex Hook. Scale bars. (A–D) = 50 μm; (E) = 25 μm.

Figure 14.
Torus. (A) Torus defined in Chamaecyparis formosensis Matsum. (SEM); (B) Torus defined in Cupressus cashmeriana Royle ex Carrière; (C) Torus not defined in Thuja plicata Donn ex D.Don. Scale bars. (A–C) = 25 μm.
Figure 14.
Torus. (A) Torus defined in Chamaecyparis formosensis Matsum. (SEM); (B) Torus defined in Cupressus cashmeriana Royle ex Carrière; (C) Torus not defined in Thuja plicata Donn ex D.Don. Scale bars. (A–C) = 25 μm.

Figure 15.
Scalloped torus and extensions. (A) Scalloped torus in Cedrus atlantica (Endl.) Manetti ex Carrière (SEM); (B) Scalloped torus in Cedrus atlantica (Endl.) Manetti ex Carrière; (C) Torus extensions in Abies magnifica A.Murray bis; (D) Torus extensions in Abies pinsapo Boiss. (SEM). Scale bars. (B,C) = 25 μm.
Figure 15.
Scalloped torus and extensions. (A) Scalloped torus in Cedrus atlantica (Endl.) Manetti ex Carrière (SEM); (B) Scalloped torus in Cedrus atlantica (Endl.) Manetti ex Carrière; (C) Torus extensions in Abies magnifica A.Murray bis; (D) Torus extensions in Abies pinsapo Boiss. (SEM). Scale bars. (B,C) = 25 μm.

Figure 16.
Notched borders. (A) Calocedrus decurrens (Torr.) Florin; (B) Athrotaxis cupressoides D.Don; (C) Taiwania cryptomerioides Hayata. Scale bars. (A–C) = 25 μm.
Figure 16.
Notched borders. (A) Calocedrus decurrens (Torr.) Florin; (B) Athrotaxis cupressoides D.Don; (C) Taiwania cryptomerioides Hayata. Scale bars. (A–C) = 25 μm.

Figure 17.
Warty layer (light microscopy). (A) Abies spectabilis (D.Don) Mirb.; (B) Callitris endlicheri (Parl.) F.M.Bailey; (C) Juniperus californica Carrière; (D) Tetraclinis articulata (Vahl) Mast. Scale bars. (A–D) = 25 μm.
Figure 17.
Warty layer (light microscopy). (A) Abies spectabilis (D.Don) Mirb.; (B) Callitris endlicheri (Parl.) F.M.Bailey; (C) Juniperus californica Carrière; (D) Tetraclinis articulata (Vahl) Mast. Scale bars. (A–D) = 25 μm.

Figure 18.
Warty layer (SEM). (A) Callitris drummondii (Parl.) Benth. and Hook.f. ex F.Muell.; (B) Callitris acuminata (Parl.) F.Muell.; (C) Callitris oblonga Rich.; (D) Diselma archeri Hook.f. Scale bars. (A,C) = 25 μm.
Figure 18.
Warty layer (SEM). (A) Callitris drummondii (Parl.) Benth. and Hook.f. ex F.Muell.; (B) Callitris acuminata (Parl.) F.Muell.; (C) Callitris oblonga Rich.; (D) Diselma archeri Hook.f. Scale bars. (A,C) = 25 μm.

Figure 19.
Organic deposits. (A,C,D) Araucaria columnaris (G.Forst.) Hook.; (B) Callitris endlicheri (Parl.) F.M.Bailey (SEM). Scale bars. (A) = 250 μm; (B,D) = 50 μm; (C) = 150 μm.
Figure 19.
Organic deposits. (A,C,D) Araucaria columnaris (G.Forst.) Hook.; (B) Callitris endlicheri (Parl.) F.M.Bailey (SEM). Scale bars. (A) = 250 μm; (B,D) = 50 μm; (C) = 150 μm.

Figure 20.
Helical thickenings. (A) Single in Taxus baccata L.; (B) In earlywood of Pseudotsuga menziesii (Mirb.) Franco; (C) In latewood of Larix decidua Mill.; (D) Grouped in pairs in Amentotaxus argotaenia (Hance) Pilg.; (E) Grouped in pairs and triplets in Torreya nucifera (L.) Siebold and Zucc.; (F) Single in Cephalotaxus harringtonia (Knight ex J.Forbes) K.Koch. Scale bars. (A) = 100 μm; (B–F) = 50 μm.
Figure 20.
Helical thickenings. (A) Single in Taxus baccata L.; (B) In earlywood of Pseudotsuga menziesii (Mirb.) Franco; (C) In latewood of Larix decidua Mill.; (D) Grouped in pairs in Amentotaxus argotaenia (Hance) Pilg.; (E) Grouped in pairs and triplets in Torreya nucifera (L.) Siebold and Zucc.; (F) Single in Cephalotaxus harringtonia (Knight ex J.Forbes) K.Koch. Scale bars. (A) = 100 μm; (B–F) = 50 μm.

Figure 21.
Helical thickenings and spiral grooves. (A) Helical thickenings in Pseudotsuga sinensis Dode. (SEM); (B) Helical thickenings in Torreya nucifera (L.) Siebold and Zucc. (SEM); (C) Spiral grooves in Neocallitropsis pancheri (Carrière) de Laub.; (D) Spiral grooves in Pinus roxburghii Sarg. (SEM); (E) Cavities caused by soft-rot in Pinus sylvestris L. resembling spiral grooves (SEM). Scale bars. (C) = 150 μm.
Figure 21.
Helical thickenings and spiral grooves. (A) Helical thickenings in Pseudotsuga sinensis Dode. (SEM); (B) Helical thickenings in Torreya nucifera (L.) Siebold and Zucc. (SEM); (C) Spiral grooves in Neocallitropsis pancheri (Carrière) de Laub.; (D) Spiral grooves in Pinus roxburghii Sarg. (SEM); (E) Cavities caused by soft-rot in Pinus sylvestris L. resembling spiral grooves (SEM). Scale bars. (C) = 150 μm.

Figure 22.
Callitroid thickenings. (A) In axial tracheids (radial section) in Callitris columellaris F.Muell.; (B,C) In axial tracheids (tangential section) in Callitris endlicheri (Parl.) F.M.Bailey; (D) In cross-fields (radial section) in Callitris columellaris F.Muell. Scale bars. (A,C,D) = 50 μm; (B) = 100 μm.
Figure 22.
Callitroid thickenings. (A) In axial tracheids (radial section) in Callitris columellaris F.Muell.; (B,C) In axial tracheids (tangential section) in Callitris endlicheri (Parl.) F.M.Bailey; (D) In cross-fields (radial section) in Callitris columellaris F.Muell. Scale bars. (A,C,D) = 50 μm; (B) = 100 μm.

Figure 23.
Callitroid thickenings (SEM). (A) In axial tracheids (radial section) in Callitris verrucosa (A.Cunn. ex Endl.) F.Muell.; (B) In axial tracheids (radial section) in Callitris endlicheri (Parl.) F.M.Bailey; (C) In cross-field pits (radial section) in Callitris verrucosa (A.Cunn. ex Endl.) F.Muell. Scale bars. (A,C) = 25 μm; (B) = 10 μm.
Figure 23.
Callitroid thickenings (SEM). (A) In axial tracheids (radial section) in Callitris verrucosa (A.Cunn. ex Endl.) F.Muell.; (B) In axial tracheids (radial section) in Callitris endlicheri (Parl.) F.M.Bailey; (C) In cross-field pits (radial section) in Callitris verrucosa (A.Cunn. ex Endl.) F.Muell. Scale bars. (A,C) = 25 μm; (B) = 10 μm.

Figure 24.
Bars of Sanio. (A) In uniseriate pits of Pinus elliottii Engelm.; (B) In biseriate pits in opposite arrangement in Pinus montezumae Lamb.; (C) In biseriate pits in opposite arrangement in Neocallitropsis pancheri (Carrière) de Laub. (SEM). Scale bars. (A,C) = 25 μm; (B) = 10 μm.
Figure 24.
Bars of Sanio. (A) In uniseriate pits of Pinus elliottii Engelm.; (B) In biseriate pits in opposite arrangement in Pinus montezumae Lamb.; (C) In biseriate pits in opposite arrangement in Neocallitropsis pancheri (Carrière) de Laub. (SEM). Scale bars. (A,C) = 25 μm; (B) = 10 μm.

Figure 25.
Trabeculae. (A) Widdringtonia nodiflora (L.) Powrie; (B) Actinostrobus pyramidalis Miq. (SEM); (C) Metasequoia glyptostroboides Hu and W.C.Cheng (SEM). Scale bars. (A) = 50 μm.
Figure 25.
Trabeculae. (A) Widdringtonia nodiflora (L.) Powrie; (B) Actinostrobus pyramidalis Miq. (SEM); (C) Metasequoia glyptostroboides Hu and W.C.Cheng (SEM). Scale bars. (A) = 50 μm.

Figure 26.
Arrangement of axial parenchyma. (A) Diffuse in Dacrydium elatum (Roxb.) Wall. ex Hook.; (B) Tangentially zonate in Juniperus excelsa M.Bieb.; (C) Marginal in Fitzroya cupressoides (Molina) I.M.Johnst.; (D) Marginal in Hesperocyparis macrocarpa (Hartw.) Bartel (Syn.—Cupressus macrocarpa Hartw.); (E) Marginal in Juniperus chinensis L. (SEM). Scale bars. (A) = 350 μm; (B) = 600 μm; (C) = 400 μm; (D) = 25 μm.
Figure 26.
Arrangement of axial parenchyma. (A) Diffuse in Dacrydium elatum (Roxb.) Wall. ex Hook.; (B) Tangentially zonate in Juniperus excelsa M.Bieb.; (C) Marginal in Fitzroya cupressoides (Molina) I.M.Johnst.; (D) Marginal in Hesperocyparis macrocarpa (Hartw.) Bartel (Syn.—Cupressus macrocarpa Hartw.); (E) Marginal in Juniperus chinensis L. (SEM). Scale bars. (A) = 350 μm; (B) = 600 μm; (C) = 400 μm; (D) = 25 μm.

Figure 27.
Transverse end walls of axial parenchyma. (A) Smooth in Cunninghamia konishii Hayata; (B) Irregularly thickened in Juniperus canariensis Guyot and Mathou; (C) Nodular in Taxodium distichum (L.) Rich.; (D) Smooth in Callitris robusta (SEM); (E) Nodular in Juniperus brevifolia (SEM). Scale bars. (A) = 50 μm; (B) = 35 μm; (C) = 50 μm; (D) = 25 μm.
Figure 27.
Transverse end walls of axial parenchyma. (A) Smooth in Cunninghamia konishii Hayata; (B) Irregularly thickened in Juniperus canariensis Guyot and Mathou; (C) Nodular in Taxodium distichum (L.) Rich.; (D) Smooth in Callitris robusta (SEM); (E) Nodular in Juniperus brevifolia (SEM). Scale bars. (A) = 50 μm; (B) = 35 μm; (C) = 50 μm; (D) = 25 μm.

Figure 28.
Ray width. (A) Uniseriate in Abies numidica de Lannoy ex Carrière; (B) Biseriate in Cedrus atlantica (Endl.) Manetti ex Carrière; (C) Fusiform in Picea alcoquiana (H.J.Veitch ex Lindl.) Carrière. Scale bars. (A–C) = 100 μm.
Figure 28.
Ray width. (A) Uniseriate in Abies numidica de Lannoy ex Carrière; (B) Biseriate in Cedrus atlantica (Endl.) Manetti ex Carrière; (C) Fusiform in Picea alcoquiana (H.J.Veitch ex Lindl.) Carrière. Scale bars. (A–C) = 100 μm.

Figure 29.
Ray composition. (A) Heterocellular ray composed of internal rows of ray parenchyma cells and marginal tracheids in Nothotsuga longibracteata (W.C.Cheng) H.H.Hu ex C.N.Page; (B) Homocellular ray composed solely of parenchyma cells in Keteleeria davidiana (C.E.Bertrand) Beissn.; (C) Heterocellular ray composed of ray tracheids in marginal rows in Pinus arizonica Engelm.; (D) Heterocellular ray composed of ray tracheids in alternate rows, both marginal and inside the ray in Pinus arizonica Engelm.; (E) Homocellular ray composed solely of ray tracheids in Pinus armandii Franch.; (F) Homocellular ray composed solely of compound ray tracheids in Callitropsis nootkatensis (D.Don) Oerst. Scale bars. (A,B) = 50 μm; (C–F) = 25 μm.
Figure 29.
Ray composition. (A) Heterocellular ray composed of internal rows of ray parenchyma cells and marginal tracheids in Nothotsuga longibracteata (W.C.Cheng) H.H.Hu ex C.N.Page; (B) Homocellular ray composed solely of parenchyma cells in Keteleeria davidiana (C.E.Bertrand) Beissn.; (C) Heterocellular ray composed of ray tracheids in marginal rows in Pinus arizonica Engelm.; (D) Heterocellular ray composed of ray tracheids in alternate rows, both marginal and inside the ray in Pinus arizonica Engelm.; (E) Homocellular ray composed solely of ray tracheids in Pinus armandii Franch.; (F) Homocellular ray composed solely of compound ray tracheids in Callitropsis nootkatensis (D.Don) Oerst. Scale bars. (A,B) = 50 μm; (C–F) = 25 μm.

Figure 30.
Ray tracheid walls. (A) Smooth in Pinus strobus L.; (B) Dentate in Pinus sylvestris L.; (C) Reticulate in Pinus pungens Lamb; (D) With helical thickenings in Pseudotsuga sinensis Dode. Scale bars. (A–D) = 25 μm.
Figure 30.
Ray tracheid walls. (A) Smooth in Pinus strobus L.; (B) Dentate in Pinus sylvestris L.; (C) Reticulate in Pinus pungens Lamb; (D) With helical thickenings in Pseudotsuga sinensis Dode. Scale bars. (A–D) = 25 μm.

Figure 31.
Pit borders of ray tracheids. (A) Pit apertures narrow in Picea alcoquiana (H.J.Veitch ex Lindl.) Carrière; (B) Pit borders with dentate thickenings in Picea abies (L.) H.Karst.; (C) Pits with wide apertures in Larix laricina (Du Roi) K.Koch. Scale bars. (A–C) = 25 μm.
Figure 31.
Pit borders of ray tracheids. (A) Pit apertures narrow in Picea alcoquiana (H.J.Veitch ex Lindl.) Carrière; (B) Pit borders with dentate thickenings in Picea abies (L.) H.Karst.; (C) Pits with wide apertures in Larix laricina (Du Roi) K.Koch. Scale bars. (A–C) = 25 μm.

Figure 32.
Horizontal walls of ray parenchyma cells. (A) Smooth in Taxodium distichum (L.) Rich.; (B) Pitted in Abies procera Rehder; (C) Pitted in Abies veitchii Lindl. (SEM); (D) Pitted in Abies cilicica (Antoine & Kotschy) Carrière (SEM). Scale bars. (A,B) = 50 μm; (D) = 25 μm.
Figure 32.
Horizontal walls of ray parenchyma cells. (A) Smooth in Taxodium distichum (L.) Rich.; (B) Pitted in Abies procera Rehder; (C) Pitted in Abies veitchii Lindl. (SEM); (D) Pitted in Abies cilicica (Antoine & Kotschy) Carrière (SEM). Scale bars. (A,B) = 50 μm; (D) = 25 μm.

Figure 33.
Nodular end walls of ray parenchyma cells. (A) Abies alba Mill.; (B) Abies cilicica (Antoine & Kotschy) Carrière (SEM); (C) Fitzroya cupressoides (Molina) I.M.Johnst.; (D) Juniperus californica Carrière (SEM). Scale bars. (A–C) = 25 μm.
Figure 33.
Nodular end walls of ray parenchyma cells. (A) Abies alba Mill.; (B) Abies cilicica (Antoine & Kotschy) Carrière (SEM); (C) Fitzroya cupressoides (Molina) I.M.Johnst.; (D) Juniperus californica Carrière (SEM). Scale bars. (A–C) = 25 μm.

Figure 34.
Indentures. (A) Calocedrus decurrens (Torr.) Florin; (B) Abies cilicica (Antoine & Kotschy) Carrière; (C) Abies lasiocarpa (Hook.) Nutt. (SEM). Scale bars. (A–C) = 25 μm.
Figure 34.
Indentures. (A) Calocedrus decurrens (Torr.) Florin; (B) Abies cilicica (Antoine & Kotschy) Carrière; (C) Abies lasiocarpa (Hook.) Nutt. (SEM). Scale bars. (A–C) = 25 μm.

Figure 35.
Irregular parenchyma cells in the marginal rows of the rays. (A) Abies fraseri (Pursh) Poir.; (B) Abies hickelii Flous and Gaussen; (C) Abies nephrolepis (Trautv.) Maxim. Scale bars. (A) = 50 μm; (B,C) = 25 μm.
Figure 35.
Irregular parenchyma cells in the marginal rows of the rays. (A) Abies fraseri (Pursh) Poir.; (B) Abies hickelii Flous and Gaussen; (C) Abies nephrolepis (Trautv.) Maxim. Scale bars. (A) = 50 μm; (B,C) = 25 μm.

Figure 36.
Cross-field pitting. (A) Window-like in Pinus resinosa Aiton; (B) Pinoid in Pinus arizonica Engelm.; (C) Piceoid in Abies cilicica (Antoine & Kotschy) Carrière; (D) Cupressoid in Juniperus canariensis, Guyot and Mathou; (E) Taxodioid in Metasequoia glyptostroboides, Hu and W.C.Cheng; (F) Araucarioid in Araucaria columnaris (G.Forst.) Hook. Scale bars. (A–F) = 25 μm.
Figure 36.
Cross-field pitting. (A) Window-like in Pinus resinosa Aiton; (B) Pinoid in Pinus arizonica Engelm.; (C) Piceoid in Abies cilicica (Antoine & Kotschy) Carrière; (D) Cupressoid in Juniperus canariensis, Guyot and Mathou; (E) Taxodioid in Metasequoia glyptostroboides, Hu and W.C.Cheng; (F) Araucarioid in Araucaria columnaris (G.Forst.) Hook. Scale bars. (A–F) = 25 μm.

Figure 37.
Cross-field pitting (SEM). (A) Window-like in Sciadopitys verticillata (Thunb.) Siebold and Zucc.; (B) Pinoid in Pinus pinea L.; (C) Piceoid in Cedrus atlantica (Endl.) Manetti ex Carrière; (D) Cupressoid in Juniperus chinensis L.; (E) Taxodioid in Glyptostrobus pensilis (Staunton ex D.Don) K.Koch; (F) Araucarioid in Araucaria cunninghamii Mudie.
Figure 37.
Cross-field pitting (SEM). (A) Window-like in Sciadopitys verticillata (Thunb.) Siebold and Zucc.; (B) Pinoid in Pinus pinea L.; (C) Piceoid in Cedrus atlantica (Endl.) Manetti ex Carrière; (D) Cupressoid in Juniperus chinensis L.; (E) Taxodioid in Glyptostrobus pensilis (Staunton ex D.Don) K.Koch; (F) Araucarioid in Araucaria cunninghamii Mudie.

Figure 38.
Resin canal subsidiary cells. (A,B) Parenchyma cells with starch content in Pinus canariensis C.Sm. ex DC.; (C,D) Strand tracheids in Picea glehnii (F.Schmidt) Mast. Scale bars. (A) = 350 μm; (B–D) = 100 μm.
Figure 38.
Resin canal subsidiary cells. (A,B) Parenchyma cells with starch content in Pinus canariensis C.Sm. ex DC.; (C,D) Strand tracheids in Picea glehnii (F.Schmidt) Mast. Scale bars. (A) = 350 μm; (B–D) = 100 μm.

Figure 39.
Normal axial resin canals. (A,B) Thick-walled epithelial cells in axial resin canals of Larix decidua Mill.; (C) Thin-walled epithelial cells in axial resin canals of Pinus radiata D.Don. and (D) Thin-walled epithelial cells in axial resin canals of Pinus sylvestris L. Scale bars. (A) = 500 μm; (B) = 100 μm; (C) = 200 μm, (D) = 100 μm.
Figure 39.
Normal axial resin canals. (A,B) Thick-walled epithelial cells in axial resin canals of Larix decidua Mill.; (C) Thin-walled epithelial cells in axial resin canals of Pinus radiata D.Don. and (D) Thin-walled epithelial cells in axial resin canals of Pinus sylvestris L. Scale bars. (A) = 500 μm; (B) = 100 μm; (C) = 200 μm, (D) = 100 μm.

Figure 40.
Normal axial and radial resin canals. (A,B) Axial resin canals in Keteleeria davidiana (C.E.Bertrand) Beissn.; (C) Thick-walled epithelial cells in radial resin canals of Picea glehnii (F.Schmidt) Mast.; (D) Thin-walled epithelial cells in radial resin canals of Pinus armandi Franch. Scale bars. (A–D) = 50 μm.
Figure 40.
Normal axial and radial resin canals. (A,B) Axial resin canals in Keteleeria davidiana (C.E.Bertrand) Beissn.; (C) Thick-walled epithelial cells in radial resin canals of Picea glehnii (F.Schmidt) Mast.; (D) Thin-walled epithelial cells in radial resin canals of Pinus armandi Franch. Scale bars. (A–D) = 50 μm.

Figure 41.
Traumatic axial and radial resin canals. (A) Traumatic resin canals in tangential rows in Keteleeria davidiana (C.E.Bertrand) Beissn.; (B) Traumatic resin canals in tangential rows in Abies nephrolepis (Trautv.) Maxim.; (C) Radial traumatic resin canals in Picea abies (L.) H.Karst. Scale bars. (A,B) = 300 μm; (C) = 150 μm.
Figure 41.
Traumatic axial and radial resin canals. (A) Traumatic resin canals in tangential rows in Keteleeria davidiana (C.E.Bertrand) Beissn.; (B) Traumatic resin canals in tangential rows in Abies nephrolepis (Trautv.) Maxim.; (C) Radial traumatic resin canals in Picea abies (L.) H.Karst. Scale bars. (A,B) = 300 μm; (C) = 150 μm.

Figure 42.
Crystals. (A) Crystals in ray parenchyma of Abies firma Siebold and Zucc.; (B) Cubic and styloid crystals in epithelial cells of axial resin canal in Pinus bungeana Zucc. ex Endl.; (C) Crystals in ray parenchyma of Abies nordmanniana subsp. equi-trojani (Asch. & Sint. ex Boiss.) Coode and Cullen. Scale bars. (A) = 25 μm; (B) = 100 μm.
Figure 42.
Crystals. (A) Crystals in ray parenchyma of Abies firma Siebold and Zucc.; (B) Cubic and styloid crystals in epithelial cells of axial resin canal in Pinus bungeana Zucc. ex Endl.; (C) Crystals in ray parenchyma of Abies nordmanniana subsp. equi-trojani (Asch. & Sint. ex Boiss.) Coode and Cullen. Scale bars. (A) = 25 μm; (B) = 100 μm.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Esteban, L.G.; de Palacios, P.; Heinz, I.; Gasson, P.; García-Iruela, A.; García-Fernández, F. Softwood Anatomy: A Review. Forests 2023, 14, 323. https://doi.org/10.3390/f14020323
AMA Style
Esteban LG, de Palacios P, Heinz I, Gasson P, García-Iruela A, García-Fernández F. Softwood Anatomy: A Review. Forests. 2023; 14(2):323. https://doi.org/10.3390/f14020323
Chicago/Turabian StyleEsteban, Luis G., Paloma de Palacios, Immo Heinz, Peter Gasson, Alberto García-Iruela, and Francisco García-Fernández. 2023. "Softwood Anatomy: A Review" Forests 14, no. 2: 323. https://doi.org/10.3390/f14020323
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.