
Altered Organic Matter Chemical Functional Groups and Bacterial Community Composition Promote Crop Yield under Integrated Soil–Crop Management System

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Soil Sampling and Analysis of Physicochemical Properties
2.3. Solid-State 13C NMR Spectroscopy
2.4. DNA Extraction and Amplicon Sequencing
2.5. Statistical Analyses
3. Results
3.1. Soil Organic Matter, Soil Fertility, and Crop Yield
3.2. Soil Organic Matter Functional Groups Depending on Management Systems
3.3. Bacterial Community Structure Depending on Management Systems
3.4. Bacterial Network and Linkage with SOM Functional Groups Depend on Management Strategy
3.5. Linking SOM Quantity and Quality, Microbial Diversity, and Soil Fertility with Crop Yield
4. Discussion
4.1. Integrated Soil–Crop Management System Increases SOM Quantity and Quality
4.2. Integrated Soil–Crop Management System Links Labile SOC with r-Strategists
4.3. SOM Quantity and Quality Increased Crop Yield by Regulating Bacterial Community Composition and Enhancing Soil Fertility
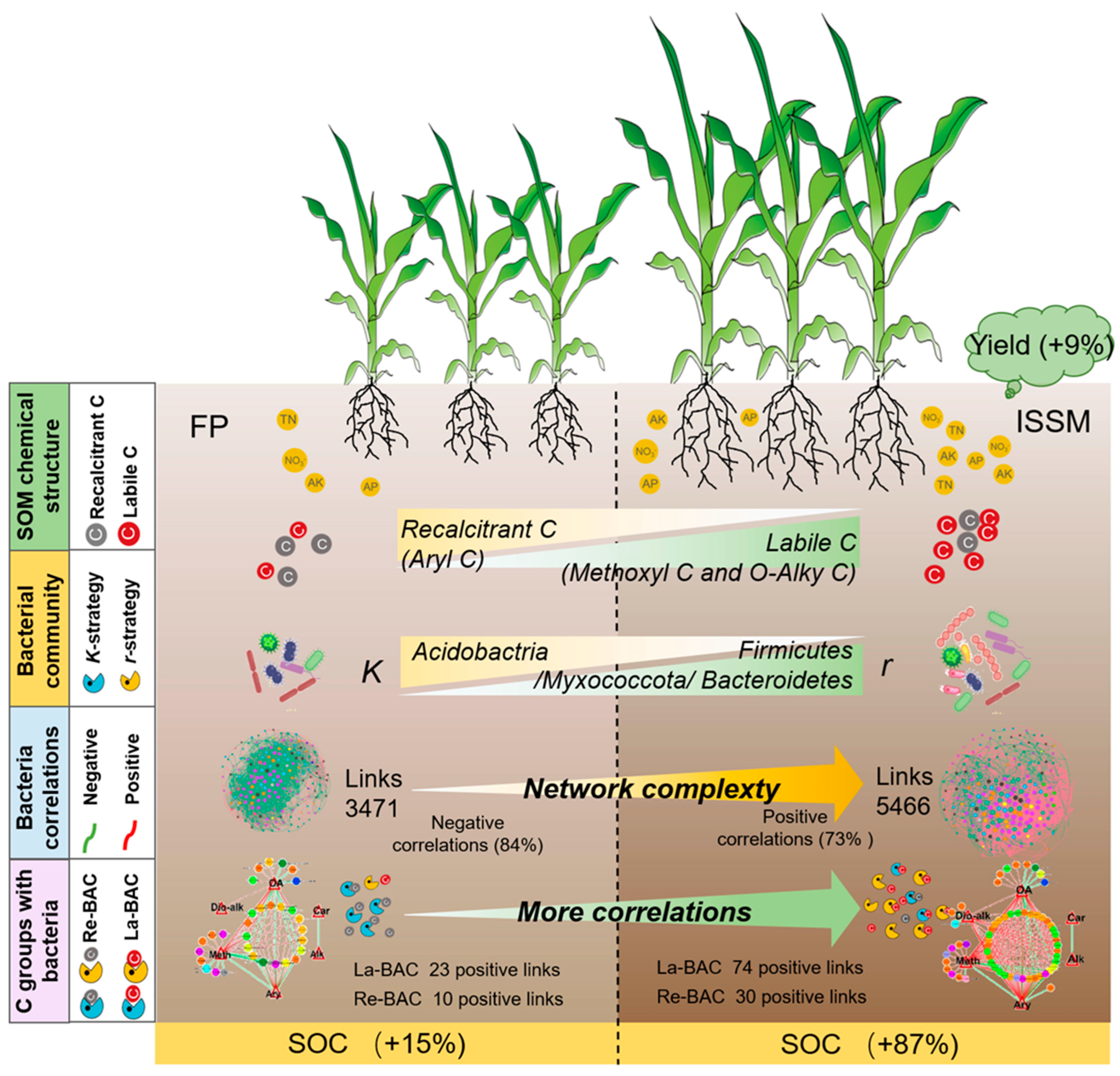
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schlesinger, W.H. Carbon Sequestration in Soils. Science 1999, 284, 2095. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, M.; Hu, S.; Zhang, X.; Ouyang, Z.; Zhang, G.; Huang, B.; Zhao, S.; Wu, J.; Xie, D.; et al. Economics- and policy-driven organic carbon input enhancement dominates soil organic carbon accumulation in Chinese croplands. Proc. Natl. Acad. Sci. USA 2018, 115, 4045–4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science 2004, 304, 1623–1627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachtergaele, F.O.; Petri, M.; Biancalani, R.; Lynden, G.V.; Velthuizen, H.V.; Bloise, M. An Information database for Land Degradation Assessment at Global Level; Sciences and Education: Bussum, The Netherland, 2011. [Google Scholar]
- Bossio, D.A.; Cook-Patton, S.C.; Ellis, P.W.; Fargione, J.; Sanderman, J.; Smith, P.; Wood, S.; Zomer, R.J.; von Unger, M.; Emmer, I.M.; et al. The role of soil carbon in natural climate solutions. Nat. Sustain. 2020, 3, 391–398. [Google Scholar] [CrossRef]
- Bradford, M.; Carey, C.; Atwood, L.; Bossio, D.; Fenichel, E.; Gennet, S.; Fargione, J.; Fisher, J.; Fuller, E.; Kane, D.; et al. Soil carbon science for policy and practice. Nat. Sustain. 2019, 2, 1070–1072. [Google Scholar] [CrossRef]
- Lal, R. Intensive Agriculture and the Soil Carbon Pool. J. Crop Improv. 2013, 27, 735–751. [Google Scholar]
- Mikha, M.M.; Rice, C.W. Tillage and Manure Effects on Soil and Aggregate-Associated Carbon and Nitrogen. Soil Sci. Soc. Am. J. 2004, 68, 809–816. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, N.; Ge, T.; Kuzyakov, Y.; Wang, Z.-L.; Li, Z.; Tang, Z.; Chen, Y.; Wu, C.; Lou, Y. Soil aggregation regulates distributions of carbon, microbial community and enzyme activities after 23-year manure amendment. Appl. Soil Ecol. 2017, 111, 65–72. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, C.; Zhang, J.; Liu, C.; Wu, Q. Effects of Substituting Manure for Fertilizer on Aggregation and Aggregate Associated Carbon and Nitrogen in a Vertisol. Agron. J. 2019, 111, 368–377. [Google Scholar] [CrossRef]
- Tian, J.; Lou, Y.; Gao, Y.; Fang, H.; Liu, S.; Xu, M.; Blagodatskaya, E.; Kuzyakov, Y. Response of soil organic matter fractions and composition of microbial community to long-term organic and mineral fertilization. Biol. Fertil. Soils 2017, 53, 523–532. [Google Scholar] [CrossRef]
- Dutta, D.; Singh, V.K.; Upadhyay, P.K.; Meena, A.L.; Kumar, A.; Mishra, R.P.; Dwivedi, B.S.; Shukla, A.K.; Yadav, G.S.; Tewari, R.B.; et al. Long-term impact of organic and inorganic fertilizers on soil organic carbon dynamics in a rice- wheat system. Land Degrad. Dev. 2022, 33, 1862–1877. [Google Scholar] [CrossRef]
- Jian, J.; Du, X.; Reiter, M.S.; Stewart, R.D. A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol. Biochem. 2020, 143, 107735. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Mei, F.; Ling, Y.; Wang, T. Does continuous straw returning keep China farmland soil organic carbon continued increase? A meta-analysis. J. Environ. Manag. 2021, 288, 112391. [Google Scholar] [CrossRef]
- De Corato, U. Agricultural waste recycling in horticultural intensive farming systems by on-farm composting and compost-based tea application improves soil quality and plant health: A review under the perspective of a circular economy. Sci. Total Environ. 2020, 738, 139840. [Google Scholar] [CrossRef]
- Fontaine, S.; Bardoux, G.; Abbadie, L.; Mariotti, A. Carbon input to soil may decrease soil carbon content. Ecol. Lett. 2010, 7, 314–320. [Google Scholar] [CrossRef]
- Luo, G.; Li, L.; Friman, V.-P.; Guo, J.; Guo, S.; Shen, Q.; Ling, N. Organic amendments increase crop yields by improving microbe-mediated soil functioning of agroecosystems: A meta-analysis. Soil Biol. Biochem. 2018, 124, 105–115. [Google Scholar] [CrossRef]
- Cai, A.; Xu, M.; Wang, B.; Zhang, W.; Liang, G.; Hou, E.; Luo, Y. Manure acts as a better fertilizer for increasing crop yields than synthetic fertilizer does by improving soil fertility. Soil Tillage Res. 2019, 189, 168–175. [Google Scholar] [CrossRef]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions, 2nd ed.; Wiley: Hoboken, NJ, USA, 1994. [Google Scholar]
- Von Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms. Soil Biol. Biochem. 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Jastrow, J.D.; Amonette, J.E.; Bailey, V.L. Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim. Change 2007, 80, 5–23. [Google Scholar] [CrossRef]
- Wang, Q.; HUANG, Q.; ZHANG, L.; ZHANG, J.; SHEN, Q.; RAN, W. The effects of compost in a rice–wheat cropping system on aggregate size, carbon and nitrogen content of the size–density fraction and chemical composition of soil organic matter, as shown by 13C CP NMR spectroscopy. Soil Use Manag. 2012, 28, 337–346. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, Z.; Zhou, H.; Wang, D.; Peng, X. The effect of 34-year continuous fertilization on the SOC physical fractions and its chemical composition in a Vertisol. Sci. Rep. 2019, 9, 2505. [Google Scholar] [PubMed] [Green Version]
- He, Y.T.; He, X.H.; Xu, M.G.; Zhang, W.J.; Yang, X.Y.; Huang, S.M. Long-term fertilization increases soil organic carbon and alters its chemical composition in three wheat-maize cropping sites across central and south China. Soil Tillage Res. 2018, 177, 79–87. [Google Scholar] [CrossRef]
- Audette, Y.; Congreves, K.A.; Schneider, K.; Zaro, G.C.; Nunes, A.L.P.; Zhang, H.; Voroney, R.P. The effect of agroecosystem management on the distribution of C functional groups in soil organic matter: A review. Biol. Fertil. Soils 2021, 57, 881–894. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Shahbaz, M.; Kuzyakov, Y.; Sanaullah, M.; Heitkamp, F.; Zelenev, V.; Kumar, A.; Blagodatskaya, E. Microbial decomposition of soil organic matter is mediated by quality and quantity of crop residues: Mechanisms and thresholds. Biol. Fertil. Soils 2017, 53, 287–301. [Google Scholar] [CrossRef]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta-analysis. Glob. Change Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, W.; Wei, W.; He, Z.; Kuzyakov, Y.; Bol, R.; Hu, R. Soil organic matter priming and carbon balance after straw addition is regulated by long-term fertilization. Soil Biol. Biochem. 2019, 135, 383–391. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Ling, L.; Luo, Y.; Jiang, B.; Lv, J.; Meng, C.; Liao, Y.; Reid, B.J.; Ding, F.; Lu, Z.; Kuzyakov, Y.; et al. Biochar induces mineralization of soil recalcitrant components by activation of biochar responsive bacteria groups. Soil Biol. Biochem. 2022, 172, 108778. [Google Scholar] [CrossRef]
- Allison, S.D.; Wallenstein, M.D.; Bradford, M.A. Soil-carbon response to warming dependent on microbial physiology. Nat. Geosci. 2010, 3, 336–340. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Ansari, M.W.; Dutta, T.; Singh, P.; Tuteja, N. Molecular characterization of cyclophilin A-like protein from Piriformospora indica for its potential role to abiotic stress tolerance in E. coli. BMC Res. Notes 2013, 6, 555. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kuzyakov, Y.; Pausch, J. Maize rhizosphere priming: Field estimates using 13C natural abundance. Plant Soil 2016, 409, 87–97. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. Organic Amendments: Microbial Community Structure, Activity and Abundance of Agriculturally Relevant Microbes Are Driven by Long-Term Fertilization Strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; Ksenofontova, N.; Zinyakova, N.B.; van Bruggen, A.H.C. Does fresh farmyard manure introduce surviving microbes into soil or activate soil-borne microbiota? J. Environ. Manag. 2021, 294, 113018. [Google Scholar] [CrossRef] [PubMed]
- Strap, J.L. Actinobacteria–Plant Interactions: A Boon to Agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 285–307. [Google Scholar]
- Bryanin, S.; Abramova, E.; Makoto, K. Fire-derived charcoal might promote fine root decomposition in boreal forests. Soil Biol. Biochem. 2018, 116, 1–3. [Google Scholar] [CrossRef]
- Li, X.; Chen, Q.L.; He, C.; Shi, Q.; Chen, S.C.; Reid, B.J.; Zhu, Y.G.; Sun, G.X. Organic Carbon Amendments Affect the Chemodiversity of Soil Dissolved Organic Matter and Its Associations with Soil Microbial Communities. Environ. Sci. Technol. 2019, 53, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.P.; Cui, Z.L.; Vitousek, P.M.; Cassman, K.G.; Matson, P.A.; Bai, J.S.; Meng, Q.F.; Hou, P.; Yue, S.C.; Romheld, V. Integrated soil–crop system management for food security. Proc. Natl. Acad. Sci. USA 2011, 108, 6399–6404. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Cui, Z.; Fan, M.; Vitousek, P.; Zhao, M.; Ma, W.; Wang, Z.; Zhang, W.; Yan, X.; Yang, J.; et al. Producing more grain with lower environmental costs. Nature 2014, 514, 486–489. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, H.; Chen, X.; Zhang, C.; Ma, W.; Huang, C.; Zhang, W.; Mi, G.; Miao, Y.; Li, X.; et al. Pursuing sustainable productivity with millions of smallholder farmers. Nature 2018, 555, 363–366. [Google Scholar] [CrossRef]
- Li, B.; Liu, Z.; Huang, F.; Yang, X.; Liu, Z.; Wan, W.; Wang, J.; Xu, Y.; Li, Z.; Ren, T. Ensuring National Food Security by Strengthening High-productivity Black Soil Granary in Northeast China. Bull. Chin. Acad. Sci. 2021, 36, 1184–1193. [Google Scholar]
- Yan, X.; CAI, Z.; WANG, S.; SMITH, P. Direct measurement of soil organic carbon content change in the croplands of China. Glob. Change Biol. 2011, 17, 1487–1496. [Google Scholar] [CrossRef]
- Liang, A.; Yang, X.; Zhang, X.; Mclaughlin, N.; Yan, S.; Li, W. Soil organic carbon changes in particle-size fractions following cultivation of Black soils in China. Soil Tillage Res. 2009, 105, 21–26. [Google Scholar] [CrossRef]
- Liu, X.; Han, X.; Song, C.; Herbert, S.J.; Xing, B. Soil Organic Carbon Dynamics in Black Soils of China Under Different Agricultural Management Systems. Commun. Soil Sci. Plant Anal. 2003, 34, 973–984. [Google Scholar] [CrossRef]
- Liu, B.; Xia, H.; Jiang, C.; Riaz, M.; Yang, L.; Chen, Y.; Fan, X.; Xia, X. 14-year applications of chemical fertilizers and crop straw effects on soil labile organic carbon fractions, enzyme activities and microbial community in rice-wheat rotation of middle China. Sci. Total Environ. 2022, 841, 156608. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.A.; Martiny, J.B.H.; Brodie, E.L.; Martiny, A.C.; Treseder, K.K.; Allison, S.D. Defining trait-based microbial strategies with consequences for soil carbon cycling under climate change. ISME J. 2020, 14, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Hanway, J.J.; Heidal, H. Soil Analysis Methods Asused in Iowa State College Soil Testing Laboratory; FAO: Rome, Italy, 1952. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Department of Agriculture: Washington, DC, USA, 1954.
- Novara, A.; Poma, I.; Sarno, M.; Venezia, G.; Gristina, L. Long-Term Durum Wheat-Based Cropping Systems Result in the Rapid Saturation of Soil Carbon in the Mediterranean Semi-arid Environment. Land Degrad. Dev. 2016, 27, 612–619. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Ma, L.; Ju, Z.; Fang, Y.; Vancov, T.; Gao, Q.; Wu, D.; Zhang, A.; Wang, Y.; Hu, C.; Wu, W.; et al. Soil warming and nitrogen addition facilitates lignin and microbial residues accrual in temperate agroecosystems. Soil Biol. Biochem. 2022, 170, 108693. [Google Scholar] [CrossRef]
- Bonanomi, G.; Incerti, G.; Giannino, F.; Mingo, A.; Lanzotti, V.; Mazzoleni, S. Litter quality assessed by solid state 13C NMR spectroscopy predicts decay rate better than C/N and Lignin/N ratios. Soil Biol. Biochem. 2013, 56, 40–48. [Google Scholar] [CrossRef]
- Baldock, J.A.; Oades, J.M.; Waters, A.G.; Peng, X.; Vassallo, A.M.; Wilson, M.A. Aspects of the chemical structure of soil organic materials as revealed by solid-state13C NMR spectroscopy. Biogeochemistry 1992, 16, 1–42. [Google Scholar] [CrossRef]
- Coxall, H.K.; Wilson, P.A.; Pälike, H.; Lear, C.H.; Backman, J. Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 2005, 433, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, P.; Chang, S.X.; Wu, J.; Lin, L. Organic mulch and fertilization affect soil carbon pools and forms under intensively managed bamboo (Phyllostachys praecox) forests in southeast China. J. Soils Sediments 2010, 10, 739–747. [Google Scholar] [CrossRef]
- Boeni, M.; Bayer, C.; Dieckow, J.; Conceição, P.C.; Dick, D.P.; Knicker, H.; Salton, J.C.; Macedo, M.C.M. Organic matter composition in density fractions of Cerrado Ferralsols as revealed by CPMAS 13C NMR: Influence of pastureland, cropland and integrated crop-livestock. Agric. Ecosyst. Environ. 2014, 190, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Cole, J.; Crowle, S.; Austwick, G.; Henderson Slater, D. Exploratory findings with virtual reality for phantom limb pain; from stump motion to agency and analgesia. Disabil. Rehabil. 2009, 31, 846–854. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Huttenhower, G.C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A new and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. On the Adaptive Control of the False Discovery Rate in Multiple Testing with Independent Statistics. J. Educ. Behav. Stat. 2000, 25, 60–83. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yao, S.; Mao, J.; Olk, D.C.; Cao, X.; Zhang, B. Chemical composition of organic matter in a deep soil changed with a positive priming effect due to glucose addition as investigated by C-13 NMR spectroscopy. Soil Biol. Biochem. 2015, 85, 137–144. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, N.; Cao, X.; Chua, T.; Mao, J.; Mandel, R.D.; Bettis, E.A.; Thompson, M.L. Composition of clay-fraction organic matter in Holocene paleosols revealed by advanced solid-state NMR spectroscopy. Geoderma 2014, 223–225, 54–61. [Google Scholar] [CrossRef]
- Arshad, M.A.; Soon, Y.K.; Ripmeester, J.A. Quality of soil organic matter and C storage as influenced by cropping systems in northwestern Alberta, Canada. Nutr. Cycl. Agroecosystems 2011, 89, 71–79. [Google Scholar] [CrossRef]
- Shrestha, B.M.; Singh, B.R.; Forte, C.; Certini, G. Long-term effects of tillage, nutrient application and crop rotation on soil organic matter quality assessed by NMR spectroscopy. Soil Use Manag. 2015, 31, 358–366. [Google Scholar] [CrossRef]
- Soon, Y.K.; Arshad, M.A.; Haq, A.; Lupwayi, N. The influence of 12 years of tillage and crop rotation on total and labile organic carbon in a sandy loam soil. Soil Tillage Res. 2007, 95, 38–46. [Google Scholar] [CrossRef]
- Li, D.; Chen, L.; Xu, J.; Ma, L.; Olk, D.C.; Zhao, B.; Zhang, J.; Xin, X. Chemical nature of soil organic carbon under different long-term fertilization regimes is coupled with changes in the bacterial community composition in a Calcaric Fluvisol. Biol. Fertil. Soils 2018, 54, 999–1012. [Google Scholar] [CrossRef]
- De Marco, A.; Spaccini, R.; Vittozzi, P.; Esposito, F.; Berg, B.; Virzo De Santo, A. Decomposition of black locust and black pine leaf litter in two coeval forest stands on Mount Vesuvius and dynamics of organic components assessed through proximate analysis and NMR spectroscopy. Soil Biol. Biochem. 2012, 51, 1–15. [Google Scholar] [CrossRef]
- Li, Y.; Chen, N.; Harmon, M.E.; Li, Y.; Cao, X.; Chappell, M.A.; Mao, J. Plant Species Rather Than Climate Greatly Alters the Temporal Pattern of Litter Chemical Composition During Long-Term Decomposition. Sci. Rep. 2015, 5, 15783. [Google Scholar] [CrossRef] [Green Version]
- Demyan, M.S.; Rasche, F.; Schulz, E.; Breulmann, M.; Müller, T.; Cadisch, G. Use of specific peaks obtained by diffuse reflectance Fourier transform mid-infrared spectroscopy to study the composition of organic matter in a Haplic Chernozem. Eur. J. Soil Sci. 2012, 63, 189–199. [Google Scholar] [CrossRef]
- Garland, G.; Edlinger, A.; Banerjee, S.; Degrune, F.; García-Palacios, P.; Pescador, D.S.; Herzog, C.; Romdhane, S.; Saghai, A.; Spor, A.; et al. Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nat. Food 2021, 2, 28–37. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Wessén, E.; Hallin, S.; Philippot, L. Differential responses of bacterial and archaeal groups at high taxonomical ranks to soil management. Soil Biol. Biochem. 2010, 42, 1759–1765. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef]
- Yuan, H.; Ge, T.; Wu, X.; Liu, S.; Tong, C.; Qin, H.; Wu, M.; Wei, W.; Wu, J. Long-term field fertilization alters the diversity of autotrophic bacteria based on the ribulose-1,5-biphosphate carboxylase/oxygenase (RubisCO) large-subunit genes in paddy soil. Appl. Microbiol. Biotechnol. 2012, 95, 1061–1071. [Google Scholar] [CrossRef]
- Bergkemper, F.; Schöler, A.; Engel, M.; Lang, F.; Krüger, J.; Schloter, M.; Schulz, S. Phosphorus depletion in forest soils shapes bacterial communities towards phosphorus recycling systems. Environ. Microbiol. 2016, 18, 1988–2000. [Google Scholar] [CrossRef] [Green Version]
- De Vries, F.T.; Bloem, J.; Quirk, H.; Stevens, C.J.; Bol, R.; Bardgett, R.D. Extensive management promotes plant and microbial nitrogen retention in temperate grassland. PLoS ONE 2012, 7, e51201. [Google Scholar] [CrossRef] [Green Version]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Kirkby, C.A.; Schmutter, D.; Bissett, A.; Kirkegaard, J.A.; Richardson, A.E. Network analysis reveals functional redundancy and keystone taxa amongst bacterial and fungal communities during organic matter decomposition in an arable soil. Soil Biol. Biochem. 2016, 97, 188–198. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.; Zhang, Y.; Zhou, J. Climate warming enhances microbial network complexity and stability. Nat. Clim. Change 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Change Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Jing, Z.; Chen, R.; Wei, S.; Feng, Y.; Zhang, J.; Lin, X. Response and feedback of C mineralization to P availability driven by soil microorganisms. Soil Biol. Biochem. 2017, 105, 111–120. [Google Scholar] [CrossRef]
- Lal, R.; Negassa, W.; Lorenz, K. Carbon sequestration in soil. Curr. Opin. Environ. Sustain. 2015, 15, 79–86. [Google Scholar] [CrossRef]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability of agricultural soils. Plant Soil 1995, 170, 75–86. [Google Scholar] [CrossRef]
- Sharma, S.K.; Ramesh, A.; Sharma, M.P.; Joshi, O.P.; Karlen, D.L. Microbial Community Structure and Diversity as Indicators for Evaluating Soil Quality. In Biodiversity, Biofuels, Agroforestry and Conservation Agriculture; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Steiner, C.; Teixeira, W.G.; Lehmann, J.; Nehls, T.; de Macêdo, J.L.V.; Blum, W.E.H.; Zech, W. Long term effects of manure, charcoal and mineral fertilization on crop production and fertility on a highly weathered Central Amazonian upland soil. Plant Soil 2007, 291, 275–290. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Sharma, P.K. Long-term effects of organics, fertilizers and cropping systems on soil physical productivity evaluated using a single value index (NLWR). Soil Tillage Res. 2008, 98, 1–10. [Google Scholar] [CrossRef]
- Sun, R.; Zhang, X.-X.; Guo, X.; Wang, D.; Chu, H. Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 2015, 88, 9–18. [Google Scholar] [CrossRef]
- Liu, H.; Du, X.; Li, Y.; Han, X.; Li, B.; Zhang, X.; Li, Q.; Liang, W. Organic substitutions improve soil quality and maize yield through increasing soil microbial diversity. J. Clean. Prod. 2022, 347, 131323. [Google Scholar] [CrossRef]
- Mazzilli, S.R.; Kemanian, A.R.; Ernst, O.R.; Jackson, R.B.; Piñeiro, G. Priming of soil organic carbon decomposition induced by corn compared to soybean crops. Soil Biol. Biochem. 2014, 75, 273–281. [Google Scholar] [CrossRef]
- Xie, H.; Li, J.; Zhu, P.; Peng, C.; Wang, J.; He, H.; Zhang, X. Long-term manure amendments enhance neutral sugar accumulation in bulk soil and particulate organic matter in a Mollisol. Soil Biol. Biochem. 2014, 78, 45–53. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Management Systems | N (kg·ha−1) | P2O5 (kg·ha−1) | K2O (kg·ha−1) | Organic Fertilizers (kg·ha−1) | Straw Management | Planting Density (Plants·ha−1) |
|---|---|---|---|---|---|---|
| Farmer practice (FP) | 300 | 120 | 120 | - | 30% | 50,000 |
| Improved farmer practice (IP) | 195 | 90 | 90 | - | 30% | 60,000 |
| High-yield system (HY) | 300 | 120 | 120 | - | 30% | 70,000 |
| Integrated soil–crop system management (ISSM) | 195 | 90 | 90 | 10,000 | 30% | 70,000 |
| Management Systems | pH | TN (g/kg) | SOC/TN | AP (mg kg−1) | AK (mg kg−1) | NO3− (mg N kg−1) | NH4+ (mg N kg−1) | BD (g m3) |
|---|---|---|---|---|---|---|---|---|
| FP | 5.04 ± 0.07 b | 1.58 ± 0.10 b | 10.6 ± 0.46 a | 85.7 ± 3.08 b | 223 ± 13.5 c | 3.57 ± 0.39 b | 6.32 ± 0.49 bc | 1.23 ± 0.00 a |
| IP | 5.27 ± 0.22 ab | 1.69 ± 0.03 b | 11.4 ± 0.31 a | 84.6 ± 3.76 b | 241 ± 16.1 bc | 14.4 ± 0.59 a | 8.94 ± 0.44 a | 1.21 ± 0.03 a |
| HY | 5.02 ± 0.12 b | 1.96 ± 0.06 b | 11.4 ± 0.31 a | 105 ± 3.99 b | 309 ± 8.35 b | 11.1 ± 1.28 a | 7.79 ± 0.38 ab | 1.18 ± 0.01 a |
| ISSM | 5.70 ± 0.10 a | 2.87 ± 0.24 a | 11.0 ± 0.59 a | 175 ± 20.7 a | 457 ± 18.8 a | 14.8 ± 0.69 a | 5.22 ± 0.55 c | 1.16 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Kumar, A.; Song, Z.; Gao, Q.; Kuzyakov, Y.; Tian, J.; Zhang, F. Altered Organic Matter Chemical Functional Groups and Bacterial Community Composition Promote Crop Yield under Integrated Soil–Crop Management System. Agriculture 2023, 13, 134. https://doi.org/10.3390/agriculture13010134
Li Q, Kumar A, Song Z, Gao Q, Kuzyakov Y, Tian J, Zhang F. Altered Organic Matter Chemical Functional Groups and Bacterial Community Composition Promote Crop Yield under Integrated Soil–Crop Management System. Agriculture. 2023; 13(1):134. https://doi.org/10.3390/agriculture13010134
Chicago/Turabian StyleLi, Qi, Amit Kumar, Zhenwei Song, Qiang Gao, Yakov Kuzyakov, Jing Tian, and Fusuo Zhang. 2023. "Altered Organic Matter Chemical Functional Groups and Bacterial Community Composition Promote Crop Yield under Integrated Soil–Crop Management System" Agriculture 13, no. 1: 134. https://doi.org/10.3390/agriculture13010134