
Microbial Biofilm Colonizing Plastic Substrates in the Ross Sea (Antarctica): First Overview of Community-Level Physiological Profiles
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
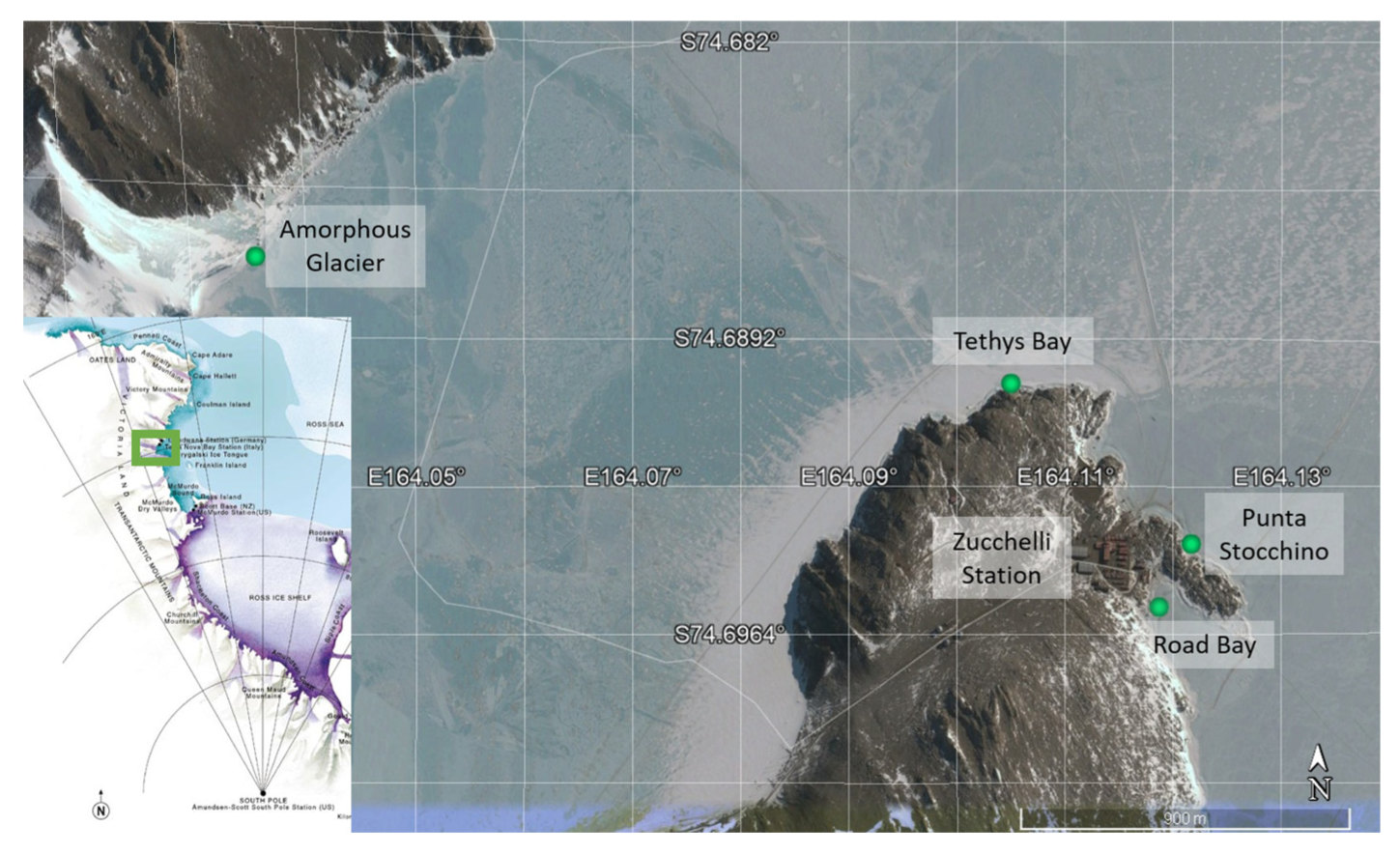
2.1. Colonization Experiment
2.2. Experimental Design
2.3. Sample Treatment
2.4. Carbon Substrate Utilization Patterns of the Microbial Biofilm Community
2.5. Statistical Analysis
3. Results
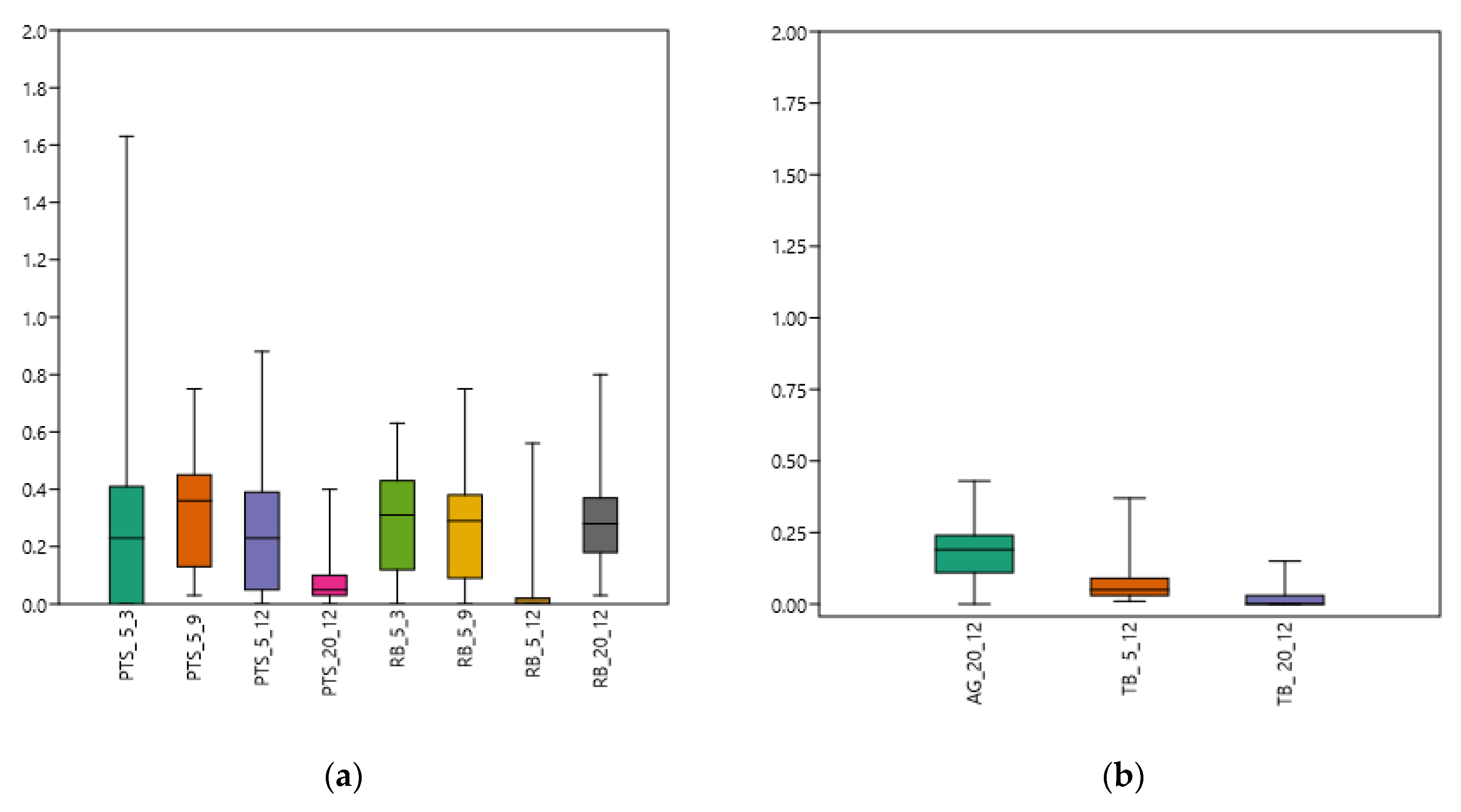
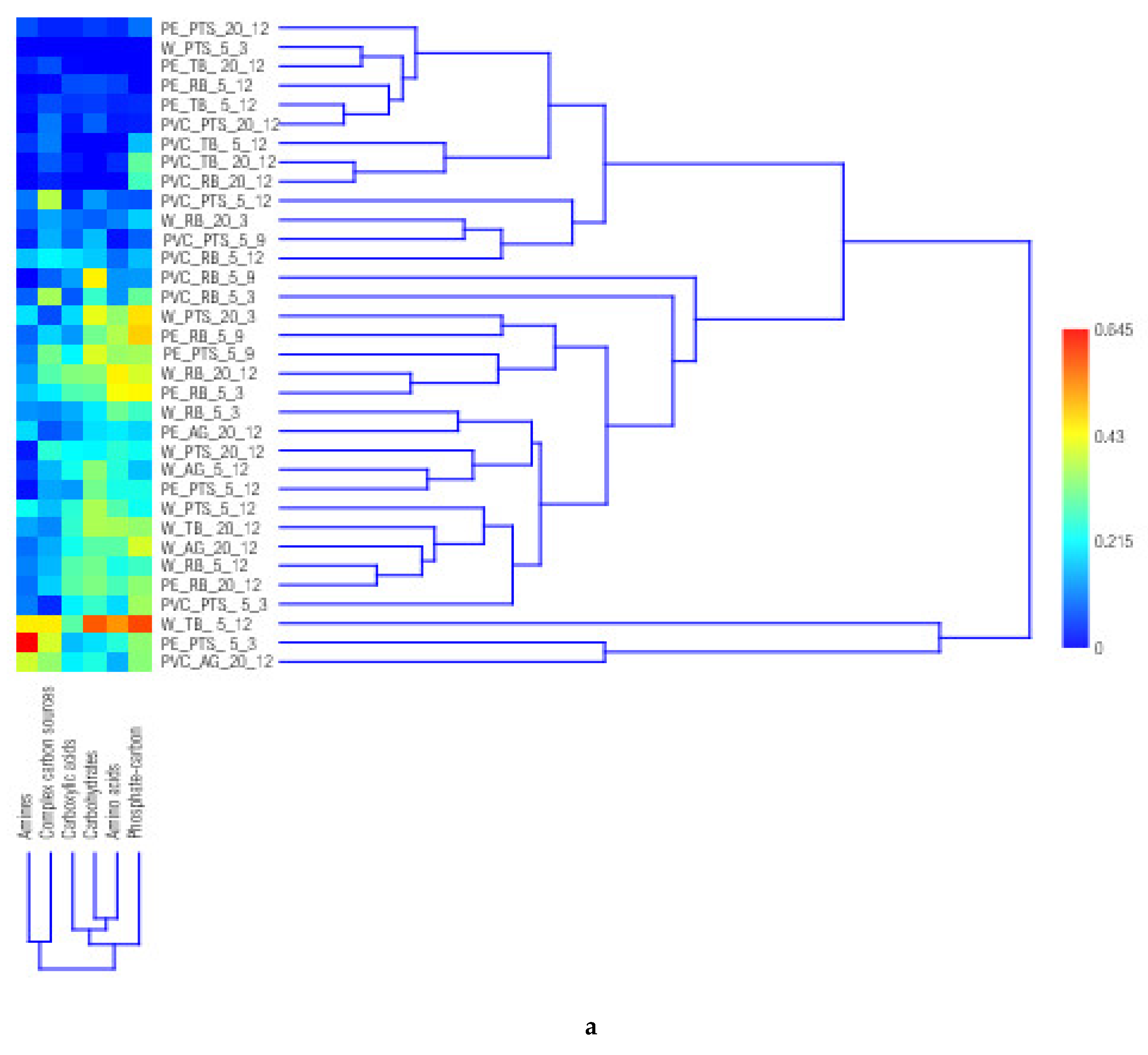
3.1. Carbon Substrate Utilization Patterns in Microbial Biofilm Community
3.2. Microbial Community on PVC Panels
3.2.1. Spatial Variability in Metabolic Utilization Rates
3.2.2. Time Variability in Metabolic Utilization Rates
3.3. Microbial Community on PE Panels
3.3.1. Spatial Variability in Metabolic Utilization Rates
3.3.2. Time Variability in Metabolic Utilization Rates
3.4. Water Samples
3.4.1. Spatial Variability in Metabolic Utilization Rates
3.4.2. Time Variability in Metabolic Utilization Rates
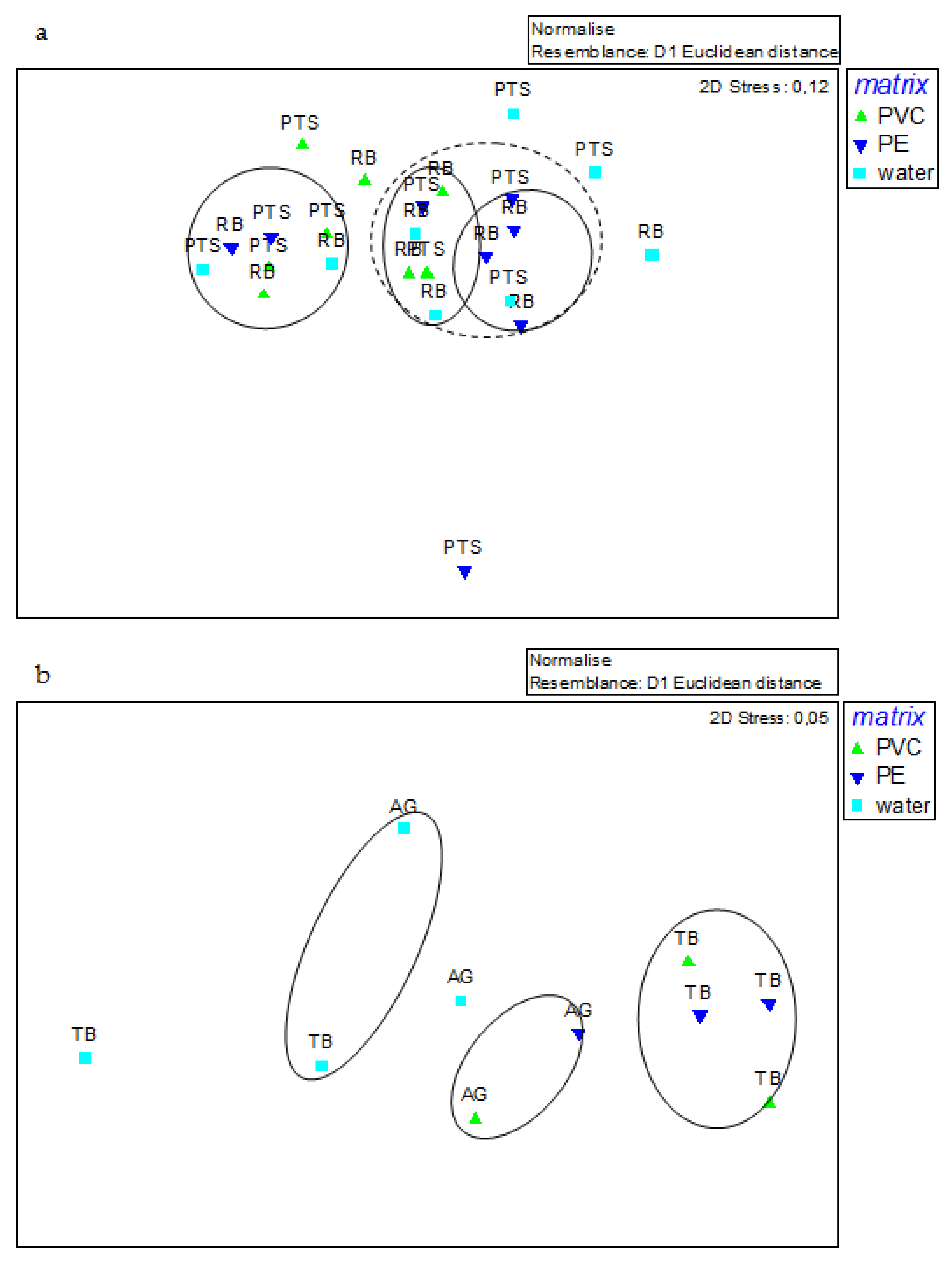
3.5. Statistical Analysis
4. Discussion
4.1. Comparison between Sampling Stations and among Matrices
4.2. Variability in Community Metabolic Profiles over a Temporal Scale
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.-F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013, 15, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.L.S.; Galdino, A.C.M.; de Mello, T.P.; de Ramos, L.S.; Branquinha, M.H.; Bolognese, A.M.; Neto, J.C.; Roudbary, M. What are the advantages of living in a community? A microbial biofilm perspective! Mem. Inst. Oswaldo Cruz 2018, 113, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef]
- Suaria, G.; Perold, V.; Lee, J.R.; Lebouard, F.; Aliani, S.; Ryan, P.G. Floating macro- and microplastics around the Southern Ocean: Results from the Antarctic Circumnavigation Expedition. Environ. Int. 2020, 136, 105494. [Google Scholar] [CrossRef] [PubMed]
- Mammo, F.K.; Amoah, I.D.; Gani, K.M.; Pillay, L.; Ratha, S.K.; Bux, F.; Kumari, S. Microplastics in the environment: Interactions with microbes and chemical contaminants. Sci. Total Environ. 2020, 743, 140518. [Google Scholar] [CrossRef]
- Corsi, I.; Bergami, E.; Caruso, G. Plastics in polar regions. Environ. Int. 2021, 149, 106203. [Google Scholar] [CrossRef]
- Caruso, G. Microbial Colonization in Marine Environments: Overview of Current Knowledge and Emerging Research Topics. J. Mar. Sci. Eng. 2020, 8, 78. [Google Scholar] [CrossRef]
- Caruso, G.; Bergami, E.; Singh, N.; Corsi, I. Plastic Occurrence, Sources, and Impacts in Antarctic Environment and Biota. Water Biol. Secur. 2022, 1, 100034. [Google Scholar] [CrossRef]
- Bargagli, R.; Rota, E. Microplastic interactions and possible combined biological effects in Antarctic marine ecosystems. Animals 2023, 3, 162. [Google Scholar] [CrossRef]
- Caruso, G. Microplastics as vectors of contaminants. Mar. Pollut. Bull. 2019, 146, 921–924. [Google Scholar] [CrossRef]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- Harrison, J.P.; Hoellein, T.J.; Sapp, M.; Tagg, A.S.; Ju-Nam, Y.; Ojeda, J.J. Microplastic-Associated Biofilms: A Comparison of Freshwater and Marine Environments. In Handbook of Environmental Chemistry; Barcelò, D., Kostianoy, A.G., Eds.; Springer: Cham, Germany, 2018; Volume 58, pp. 181–201. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Labrenz, M. Marine Microbial Assemblages on Microplastics: Diversity, Adaptation, and Role in Degradation. Ann. Rev. Mar. Sci. 2020, 12, 209–232. [Google Scholar] [CrossRef]
- Pinto, M.; Langer, T.M.; Hüer, T.; Hofmann, T.; Herndl, G.J. The composition of bacterial communities associated with plastic biofilms differs between different polymers and stages of biofilm succession. PLoS ONE 2019, 14, e0217165. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, M.; Collard, F.; Fabres, J.; Gabrielsen, G.W.; Provencher, J.F.; Rochman, C.M.; van Sebille, E.; Tekman, M.B. Plastic pollution in the Arctic. Nat. Rev. Earth Environ. 2022, 3, 323–337. [Google Scholar] [CrossRef]
- Webster, N.S.; Negri, A.P. Site-specific variation in Antarctic marine biofilms established on artificial surfaces. Environ. Microbiol. 2006, 8, 1177–1190. [Google Scholar] [CrossRef]
- Lee, Y.M.; Cho, K.H.; Hwang, K.; Kim, E.H.; Kim, M.; Hong, S.G.; Lee, H.K. Succession of bacterial community structure during the early stage of biofilm development in the Antarctic marine environment. Korean J. Microbiol. 2016, 52, 49–58. [Google Scholar] [CrossRef]
- Clark, M.S.; Villota Nieva, L.; Hoffman, J.I.; Davies, A.J.; Trivedi, U.J.; Turner, F.; Ashton, G.V.; Peck, L.S. Lack of long-term acclimation in Antarctic encrusting species suggests vulnerability to warming. Nat. Commun. 2019, 10, 3383. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.R.; Slikas, B.; Boyd, G.D.; Melvin, D.W.; Morrall, C.E.; Proskurowski, G.; Mincer, T.J. The biogeography of the Plastisphere: Implications for policy. Front. Ecol. Environ. 2015, 13, 541–546. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J. Ecology of the plastisphere. Nat. Rev. Microbiol. 2020, 18, 139–151. [Google Scholar] [CrossRef]
- Cappello, S.; Caruso, G.; Bergami, E.; Macrì, A.; Venuti, V.; Majolino, D.; Corsi, I. New insights into the structure and function of the prokaryotic communities colonizing plastic debris collected in King George Island (Antarctica): Preliminary observations from two plastic fragments. J. Hazard. Mat. 2021, 414, 125586. [Google Scholar] [CrossRef]
- Cavicchioli, R. Microbial ecology of Antarctic aquatic systems. Nat. Rev. Microbiol. 2015, 13, 691–706. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.O., Jr.; Sedwick, P.N.; Arrigo, K.R.; Ainley, D.G.; Orsi, A.H. The Ross Sea in a sea of change. Oceanography 2012, 25, 90–103. [Google Scholar] [CrossRef]
- Fabiano, M.; Danovaro, R. Enzymatic Activity, Bacterial Distribution, and Organic Matter Composition in Sediments of the Ross Sea (Antarctica). Appl. Environ. Microbiol. 1998, 64, 3838–3845. [Google Scholar] [CrossRef] [PubMed]
- Celussi, M.; Cataletto, B.; Fonda Umani, S.; Del Negro, P. Depth profiles of bacterioplankton assemblages and their activities in the Ross Sea. Deep Sea Res. Part I 2009, 56, 2193–2205. [Google Scholar] [CrossRef]
- Celussi, M.; Bergamasco, A.; Cataletto, B.; Umani, S.F.; Del Negro, P. Water masses’ bacterial community structure and microbial activities in the Ross Sea, Antarctica. Antarct. Sci. 2010, 22, 361–370. [Google Scholar] [CrossRef]
- Azzaro, M.; Specchiulli, A.; Maimone, G.; Azzaro, F.; Lo Giudice, A.; Papale, M.; La Ferla, R.; Paranhos, R.; Souza Cabral, A.; Rappazzo, A.C.; et al. Trophic and Microbial Patterns in the Ross Sea Area (Antarctica): Spatial Variability during the Summer Season. J. Mar. Sci. Eng. 2022, 10, 1666. [Google Scholar] [CrossRef]
- Cummings, V.J.; Hewitt, J.E.; Thrush, S.F.; Marriott, P.M.; Halliday, N.J.; Norkko, A. Linking Ross Sea Coastal Benthic Communities to Environmental Conditions: Documenting Baselines in a Spatially Variable and Changing World. Front. Mar. Sci. 2018, 5, 232. [Google Scholar] [CrossRef]
- Ylla, I.; Canhoto, C.; Romaní, A.M. Effects of warming on stream biofilm organic matter use capabilities. Microb. Ecol. 2014, 68, 132–145. [Google Scholar] [CrossRef]
- Romaní, A.M.; Borrego, C.M.; Diaz-Villanueva, V.; Freixa, A.; Gich, F.; Ylla, I. Shifts in microbial community structure and function in light- and dark-grown biofilms driven by warming. Environ. Microbiol. 2014, 16, 2550–2567. [Google Scholar] [CrossRef]
- La Ferla, R.; Azzaro, M.; Michaud, L.; Caruso, G.; Lo Giudice, A.; Paranhos, R.; Cabral, A.S.; Conte, A.; Cosenza, A.; Maimone, G.; et al. Prokaryotic Abundance and Activity in Permafrost of the Northern Victoria Land and Upper Victoria Valley (Antarctica). Microb. Ecol. 2017, 74, 402–415. [Google Scholar] [CrossRef]
- Papale, M.; Lo Giudice, A.; Conte, A.; Rizzo, C.; Rappazzo, A.C.; Maimone, G.; Caruso, G.; La Ferla, R.; Azzaro, M.; Gugliandolo, C.; et al. Microbial Assemblages in Pressurized Antarctic Brine Pockets (Tarn Flat, Northern Victoria Land): A Hotspot of Biodiversity and Activity. Microorganisms 2019, 7, 333. [Google Scholar] [CrossRef]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [PubMed]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Sala, M.M.; Arin, L.; Balagué, V.; Felipe, J.; Guadayol, O.; Vaqué, D. Functional diversity of bacterioplankton assemblages in Western Antarctic seawaters during late spring. Mar. Ecol. Progr. Ser. 2005, 292, 13–21. [Google Scholar] [CrossRef]
- Sala, M.M.; Estrada, M.; Gasol, J.M. Seasonal changes in the functional diversity of bacterioplankton in contrasting coastal environments of the NW Mediterranean. Aquat. Microb. Ecol. 2006, 44, 1–9. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Plymouth, UK, 2014. [Google Scholar]
- Margalef, R. Ecología; Omega: Barcelona, Spain, 1977; pp. 1–951. [Google Scholar]
- Ludwig, J.A.; Reynolds, J.F. Statistical Ecology: A Primer on Methods and Computing; Wiley: New York, NY, USA, 1988; pp. 85–106. [Google Scholar]
- Pielou, E.C. Ecological Diversity; John Wiley & Sons: New York, NY, USA, 1975. [Google Scholar]
- Zanne, A.E.; Abarenkov, K.; Afkhami, M.E.; Aguilar-Trigueros, C.A.; Bates, S.; Bhatnagar, J.M.; Busby, P.E.; Christian, N.; Cornwell, W.K.; Crowther, T.W.; et al. Fungal functional ecology: Bringing a trait-based approach to plant-associated fungi. Biol. Rev. 2020, 95, 409–433. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. Variations in biofilms colonizing artificial surfaces: Seasonal effects and effects of grazers. J. Mar. Biol. Ass. 1995, 75, 705–714. [Google Scholar] [CrossRef]
- Caruso, G.; Pedà, C.; Cappello, S.; Leonardi, M.; La Ferla, R.; Giudice, A.L.; Maricchiolo, G.; Rizzo, C.; Maimone, G.; Rappazzo, A.C.; et al. Effects of microplastics on trophic parameters, abundance and metabolic activities of seawater and fish gut bacteria in mesocosm conditions. Environ. Sci. Pollut. Res. 2018, 25, 30067–30083. [Google Scholar] [CrossRef]
- Caroppo, C.; Azzaro, M.; Dell’Acqua, O.; Azzaro, F.; Maimone, G.; Rappazzo, A.C.; Raffa, F.; Caruso, G. Microbial Biofilms Colonizing Plastic Substrates in the Ross Sea (Antarctica). J. Mar. Sci. Eng. 2022, 10, 1714. [Google Scholar] [CrossRef]
- Caruso, G.; Dell’Acqua, O.; Caruso, R.; Azzaro, M. Phenotypic characterization of bacterial isolates from marine waters and plastisphere communities of the Ross Sea (Antarctica). J. Clin. Microbiol. Biochem. Technol. 2022, 8, 001–009. [Google Scholar] [CrossRef]
- Zhang, W.; Ding, W.; Li, Y.-X.; Tam, C.; Bougouffa, S.; Wang, R.; Pei, B.; Chiang, H.; Leung, P.; Lu, Y.; et al. Marine biofilms constitute a bank of hidden microbial diversity and functional potential. Nat. Commun. 2019, 10, 517. [Google Scholar] [CrossRef]
- Laganà, P.; Votano, L.; Caruso, G.; Azzaro, M.; Lo Giudice, A.; Delia, S. Bacterial isolates from the Arctic region (Pasvik River, Norway): Assessment of biofilm production and antibiotic susceptibility profiles. Environ. Sci. Pollut. Res. 2018, 25, 1089–1102. [Google Scholar] [CrossRef]
- Sottile, C. Evaluation of the Capacity of Producing Biofilm in Bacterial Strains from the Antarctica (In Italian). Bachelor’s Thesis, University of Messina, Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, Messina, Italy, 2020; pp. 1–61. [Google Scholar]
- Caruso, G. Microbial Colonization of Antarctic Benthic Environments: Response of Microbial Abundances, Diversity, Activities and Larval Settlement to Natural and Anthropogenic Disturbances and Search for Secondary Metabolites (ANT-Biofilm, PNRA16_00105); Final Scientific Report; Programma Nazionale di Ricerche in Antartide (PNRA): Rome, Italy, 2020; Available online: https://www.pnra.aq/en/project/335/microbial-colonization-benthic-environments-antarctica-responses-microbial-abundances (accessed on 15 March 2023).
- Oliver, J.D. Taxonomic scheme for the identification of marine bacteria. Deep Sea Res. Part A Oceanogr. Res. Pap. 1982, 29, 795–798. [Google Scholar] [CrossRef]
- Sujarit, D.; Lyla, P.; Khan, S. A simple scheme for the identification of marine heterotrophic bacteria. Thalassas 2007, 23, 17–21. [Google Scholar]
- Litchfield, C.D.; Irby, A.; Kis-Papo, T.; Oren, A. Comparative metabolic diversity in two solar salterns. Hydrobiologia 2001, 466, 73–80. [Google Scholar] [CrossRef]
- Miao, L.; Wang, C.; Adyel, T.; Zhao, J.; Yan, N.; Wu, J.; Hou, J. Periphytic biofilm formation on natural and artificial substrates: Comparison of microbial composition, interaction, and function. Front. Microbiol. 2021, 12, 1917. [Google Scholar] [CrossRef] [PubMed]
- Gilichinsky, D.; Vishnivetskaya, T.; Petrova, M. Bacteria in permafrost. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Schinner, F., Marx, J.C., Gerday, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 83–102. [Google Scholar]
- Jałowiecki, Ł.; Chojniak, J.M.; Dorgeloh, E.; Hegedusova, B.; Ejhed, H.; Magnér, J.; Płaza, G.A. Microbial community profiles in wastewaters from onsite wastewater treatment systems technology. PLoS ONE 2016, 11, e0147725. [Google Scholar] [CrossRef] [PubMed]
- Kenarova, A.; Encheva, M.; Chipeva, V. Physiological diversity of bacterial communities from different soil locations on Livingston Island, South Shetland archipelago, Antarctica. Polar Biol. 2013, 36, 223–233. [Google Scholar] [CrossRef]
- Papale, M.; Caruso, G.; Maimone, G.; La Ferla, R.; Giudice, A.L.; Rappazzo, A.C.; Cosenza, A.; Azzaro, F.; Ferretti, R.; Paranhos, R.; et al. Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle. Water 2023, 15, 556. [Google Scholar] [CrossRef]
- Pessi, I.S.; De Oliveira Elias, S.; Simoes, F.L.; Simoes, J.C.; Macedo, A.J. Functional diversity of microbial communities in soils in the vicinity of Wanda Glacier, Antarctic Peninsula. Microbes Environ. 2012, 27, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Osono, T.; Hasegawa, M.; Hobara, S.; Hirose, D.; Tanabe, Y.; Kudoh, S.; Uchida, M. Application of the Biolog EcoPlate™ technique for assessing the microbial metabolic diversity in moss profiles of continental Antarctica. Polar Sci. 2023, 35, 100924. [Google Scholar] [CrossRef]
- Sanyal, A.; Antony, R.; Samui, G.; Thamban, M. Microbial communities and their potential for degradation of dissolved organic carbon in cryoconite hole environments of Himalaya and Antarctica. Microbiol. Res. 2018, 208, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Sala, M.M.; Terrado, R.; Lovejoy, C.; Unrein, F.; Pedrớs-Aliớ, C. Metabolic diversity of heterotrophic bacterioplankton over winter and spring in the coastal Arctic Ocean. Environ. Microbiol. 2008, 10, 942–949. [Google Scholar] [CrossRef]
- Yergeau, E.; Kowalchuk, G.A. Responses of Antarctic soil microbial communities and associated functions to temperature and freeze-thaw cycle frequency. Environ. Microbiol. 2008, 10, 2223–2235. [Google Scholar] [CrossRef]
- Wu, Y.; Cai, P.; Jing, X.; Niu, X.; Ji, D.; Ashry, N.M.; Gao, C.; Huang, Q. Soil biofilm formation enhances microbial community diversity and metabolic activity. Environ. Int. 2019, 132, 105116. [Google Scholar] [CrossRef]
- Thomas, F.A.; Mohan, M.; Krishnan, K.P. Bacterial diversity and their metabolic profiles in the sedimentary environments of Ny-Ålesund, Arctic. Antonie Van Leeuwenhoek 2021, 114, 1339–1360. [Google Scholar] [CrossRef]
- A’lvarez-Añorve, L.I.; Calcagno, M.L.; Plumbridge, J. Why does Escherichia coli grow more slowly on glucosamine than on N-acetylglucosamine? Effects of enzyme levels and allosteric activation of GlcN6P deaminase (NagB) on growth rates. J. Bacteriol. 2005, 187, 2974–2982. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Moore, J.A.; Kirchman, D.L. Chitinases from Uncultured Marine Microorganisms. Am. Soc. Microbiol. 1999, 65, 2553–2557. [Google Scholar] [CrossRef]
- Reitermayer, D.; Kafka, T.A.; Lenz, C.A.; Vogel, R.F. Interrelation between Tween and the membrane properties and high pressure tolerance of Lactobacillus plantarum. BMC Microbiol. 2018, 18, 72. [Google Scholar] [CrossRef]
- Collins, T.; Margesin, R. Psychrophilic lifestyles: Mechanisms of adaptation and biotechnological tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Teske, A.; Durbin, A.; Ziervogel, K.; Cox, C.; Arnosti, C. Microbial community composition and function in permanently cold seawater and sediments from an Arctic fjord of Svalbard. Appl. Environ. Microbiol. 2011, 77, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Krishnan, K.P. Differences in free-living and particle-associated bacterial communities and their spatial variation in Kongsfjorden, Arctic. J. Basic Microbiol. 2017, 57, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gómez, B.; Sala, M.M.; Pedrós-Alió, C. Seasonal changes in substrate utilization patterns by bacterioplankton in the Amundsen Gulf (western Arctic). Polar Biol. 2014, 37, 1321–1329. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | PTS | RB | TB | AG |
|---|---|---|---|---|
| PVC | ||||
| 5 m (T3) | 48 | 48 | / | / |
| 5 m (T9) | 48 | 48 | / | / |
| 5 m (T12) | 48 | 72 | 72 | / |
| 20 m (T12) | 0 | 48 | 48 | 0/72 |
| PE | ||||
| 5 m (T3) | 48 | 336 | ||
| 5 m (T9) | 48 | 48 | / | / |
| 5 m (T12) | 48 | 48 | 72 | / |
| 20 m (T12) | 72 | 48 | 288 | 0/48 |
| Water | ||||
| 5 m (T12) | 96 | 96 | 48 | 96 |
| 20 m (T12) | 168 | 168 | 96 | 96 |
| PVC | d | S | J′ | H′ (log e) |
| PTS_ 5_3 | 13.08 | 26 | 0.9392 | 3.060 |
| PTS_5_9 | 15.62 | 25 | 0.9301 | 2.994 |
| PTS_5_12 | 15.48 | 26 | 0.9047 | 2.948 |
| PTS_20_12 | 26.02 | 24 | 0.9000 | 2.860 |
| RB_5_3 | 13.77 | 28 | 0.9472 | 3.156 |
| RB_5_9 | 13.43 | 28 | 0.8972 | 2.990 |
| RB_5_12 | 15.47 | 29 | 0.9196 | 3.096 |
| RB_20_12 | 11.42 | 13 | 0.8577 | 2.200 |
| AG_20_12 | 12.99 | 27 | 0.9594 | 3.162 |
| TB_ 5_12 | 32.40 | 18 | 0.9015 | 2.606 |
| TB_ 20_12 | 16.16 | 20 | 0.8758 | 2.624 |
| PE | d | S | J′ | H′(log e) |
| PTS_ 5_3 | 8.79 | 22 | 0.8982 | 2.776 |
| PTS_5_9 | 13.03 | 31 | 0.9433 | 3.239 |
| PTS_5_12 | 14.35 | 30 | 0.9075 | 3.086 |
| PTS_20_12 | 32.51 | 30 | 0.9098 | 3.094 |
| RB_5_3 | 12.04 | 27 | 0.9600 | 3.164 |
| RB_5_9 | 13.34 | 29 | 0.9433 | 3.176 |
| RB_5_12 | 17.26 | 8 | 0.8683 | 1.806 |
| RB_20_12 | 13.56 | 31 | 0.9464 | 3.250 |
| AG_20_12 | 16.32 | 29 | 0.9707 | 3.269 |
| TB_ 5_12 | 34.94 | 31 | 0.9005 | 3.092 |
| TB_ 20_12 | 14 | 0.8471 | 2.236 | |
| Water | d | S | J′ | H′(log e) |
| PTS_5_3 | 4 | 0.9323 | 1.292 | |
| PTS_5_12 | 12.53 | 31 | 0.9568 | 3.286 |
| PTS_20_3 | 12.03 | 27 | 0.9552 | 3.148 |
| PTS_20_12 | 12.69 | 30 | 0.9483 | 3.225 |
| RB_5_3 | 13.77 | 28 | 0.9729 | 3.242 |
| RB_5_12 | 14.94 | 31 | 0.9288 | 3.190 |
| RB_20_3 | 19.20 | 29 | 0.9595 | 3.231 |
| RB_20_12 | 12.15 | 31 | 0.9628 | 3.306 |
| AG_5_12 | 12.91 | 28 | 0.9451 | 3.149 |
| AG_20_12 | 13.67 | 31 | 0.9508 | 3.265 |
| TB_ 5_12 | 11.24 | 31 | 0.9580 | 3.290 |
| TB_ 20_12 | 12.35 | 30 | 0.9665 | 3.287 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; Maimone, G.; Rappazzo, A.C.; Dell’Acqua, O.; Laganà, P.; Azzaro, M. Microbial Biofilm Colonizing Plastic Substrates in the Ross Sea (Antarctica): First Overview of Community-Level Physiological Profiles. J. Mar. Sci. Eng. 2023, 11, 1317. https://doi.org/10.3390/jmse11071317
Caruso G, Maimone G, Rappazzo AC, Dell’Acqua O, Laganà P, Azzaro M. Microbial Biofilm Colonizing Plastic Substrates in the Ross Sea (Antarctica): First Overview of Community-Level Physiological Profiles. Journal of Marine Science and Engineering. 2023; 11(7):1317. https://doi.org/10.3390/jmse11071317
Chicago/Turabian StyleCaruso, Gabriella, Giovanna Maimone, Alessandro Ciro Rappazzo, Ombretta Dell’Acqua, Pasqualina Laganà, and Maurizio Azzaro. 2023. "Microbial Biofilm Colonizing Plastic Substrates in the Ross Sea (Antarctica): First Overview of Community-Level Physiological Profiles" Journal of Marine Science and Engineering 11, no. 7: 1317. https://doi.org/10.3390/jmse11071317