
Effect of Bed Preparation on Native Wildflower Establishment, Weed Control, and Arthropod Presence


1
HeartBound Ministries, Atlanta, GA 31119, USA
2
Department of Horticulture, University of Georgia-Griffin, 290 Higgins Rd, Griffin, GA 30223, USA
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(8), 854; https://doi.org/10.3390/horticulturae9080854
Submission received: 23 June 2023
/
Revised: 17 July 2023
/
Accepted: 24 July 2023
/
Published: 27 July 2023
(This article belongs to the Section Floriculture, Nursery and Landscape, and Turf)
Abstract
:Native flowering forbs plantings have been found to support diverse arthropod communities by providing year-round forage and refuge from prey and chemical pollutants. Typically, pollinator habitats are planted from seeds; however, poor establishment and weed pressure often result in limited success of the planting. In this study, we evaluated two bed preparation techniques, a glyphosate treatment with no tillage and tillage with soil fumigant (dazomet) to determine their impact on herbaceous perennial transplants establishment, weed control, and arthropod presence. Our results suggest that forbs grown in beds that are tilled and treated with dazomet exhibit greater growth, produce more blooms, and experience reduced weed pressure compared to plants grown in untilled beds treated with glyphosate. We also found that the bed preparation method had no effect on arthropod activity, despite higher bloom counts in plants grown in beds prepared with tillage and dazomet applications. This research indicates that of the fifteen native flowering forbs examined in this study, a majority attracted abundant and diverse insect populations, despite variations in plant growth and bloom counts due to bed preparation treatments.
1. Introduction
A variety of cultural and political solutions have been proposed to combat biodiversity declines on local, regional, and national scales in many countries across the globe [1]. Suggestions include incentivizing farmers to build pollinator habitats on private land [2], removing invasive plants [3], reducing herbicide and insecticide use [4], and planting native vegetation along roadsides [5]. Despite the high public interest, homeowners, landscaping companies, and civic entities are often deterred from planting native wildflowers because they typically require complete renovation every two to three years [6,7]. Additionally, traditional herbicides cannot be used to manage weeds [8,9,10,11], seed mixes can be costly, and a limited number of species (often a single one) tend to dominate the space over time [7], creating a monoculture lacking in arthropod and plant diversity.
Research has documented the ecological benefits of wildflowers plantings in Europe [8,12] and North America [13], among other regions of the world. Despite these benefits, however, plantings often face significant weed pressure [14], which can result in limited long-term success of the habitat. Conflicting information exists regarding the best methods for ensuring habitat longevity [15,16,17,18,19], with some studies showing better results with soil disturbance, while others point to the opposite conclusion.
A multitude of bed preparation methods have been studied, including grazing, solarization, burning, herbicide applications, and tillage [17]. Outside of organic bed preparation techniques, glyphosate has been the standard non-selective herbicide used to clear all vegetation in preparation of wildflower plantings [20]. Glyphosate targets actively growing plants [21] and is rapidly inactivated in soil [22]. Dazomet is a broad-spectrum pre-plant soil fumigant that has been used widely to control weeds and soil-borne pathogens in a wide variety of horticulture crops [23,24] We set out to investigate the effectiveness of the soil fumigant dazomet as a tool for weed suppression in tillage treatment systems compared to no-till treatment with glyphosate. In order for dazomet to be successful, it must be incorporated into the soil through tillage (4 to 6 inches), irrigated with approximately one inch of water and covered immediately with plastic to seal the fumes that are produced. The resulting volatile sterilizes the soil, thus alleviating much of the disease, weed, insect, nematode, and insect pressure (Basamid G® label). Dazomet has not been well studied as a tool for wildflower bed preparation, but is used for bed preparation in Christmas tree production [25]. While dazomet is a restricted-use pesticide [26] and not available to the public, it is a tool that is available to landscaping companies and land managers.
Most wildflower plantings are established with direct seeding [27]. Unfortunately, growing wildflowers from seeds presents a myriad of problems. Seed germination is erratic, and many species will not bloom in their first year, leaving beds with large open spaces that can be overrun by weeds [28]. Seed mix germination rates vary from year to year and company to company [29], potentially reducing diversity, which is one of the essential elements of a healthy, sustainable, pollinator-friendly wildflower planting [30]. Seed establishment is dependent on a variety of factors, including seed viability, ability to overcome dormancy, light, temperature, oxygen, moisture, planting date, and seed to soil contact [17]. Furthermore, once seeds have been laid, growers must rely on seasonal rainfall if supplemental irrigation is not available [5]. According to the Environmental Protection Agency, as the average temperature of Earth’s surface continues to rise, more evaporation occurs, increasing overall precipitation [26]. While increased irrigation might be beneficial for maintaining wildflower plantings, more rainfall might have an unintended consequence; seeds can wash away or the soil can become too saturated, thus depleting soil oxygen levels and stunting germination and growth [31]. Additionally, many wildflower beds are sown in spring, and a late frost can destroy seedlings [17].
Plugs are increasingly recommended as an alternative to seeds in native grass and wildflower plantings, yet wildflower bed establishment from plugs is not well researched. Plugs allow for quick establishment of a wildflower planting, especially species that are either slow to germinate, difficult to establish, or have expensive seeds [17]. Finally, plants transplanted as plugs bloom earlier and create an immediate green mulch effect by blocking sunlight from reaching the ground, thus preventing weed germination [32].
Although perennial forbs planted as plugs has been previously used in research, the aim of those studies was assessing arthropod attraction [33,34,35,36], and the experiment was set up as a garden under irrigated conditions [37,38]. Evaluating plant establishment from transplants under non-irrigated conditions has not received much attention.
Native plants are essential for the remediation of fragmented and limited-resource urban landscapes because they serve as sources of pollen, nectar, and shelter for pollinators and beneficial insects [39]. Urban and peri-urban habitats are considered challenging for plant growth due to factors such as soil compaction, restricted water movement [40], soil moisture extremes, and higher aerial and soil temperatures [41]. Native forbs vary in their adaptability to such environments, yet interest in using native plants as floral resources in arthropod conservation programs has increased in the last decade [42]. Screening native species for adaptability to challenging environments while assessing their potential to support pollinators and natural enemies is the basis for formulating data-driven decisions in conservation programs [33].
Arthropod communities are complex networks of ground-dwelling species and those that spend most time aboveground—all revolving around the basic concept of finding shelter, food, and mates [43]. Plants provide food for herbivorous insects (including bees, as pollen and nectar are of plant origin) and in turn, those insects attract carnivorous species. A healthy arthropod assemblage comprises numerous species [44]. Few studies have explored the entire arthropod community (i.e., flying and ground-dwelling species) when studying wildflower habitats; most studies have focused on aboveground communities. A recent study found that many, but not all, arthropod taxa profit from wildflower meadows in terms of arthropod activity, abundance in pitfall traps, and arthropod density in standardized suction samples [45]. Arthropod abundance in meadows was 212% higher in pitfall traps and 260% higher in suction samples compared to woody vegetation [45].
Soil tillage can disturb ground-dwelling insects, especially ground-nesting bee species [46]. Most solitary bee species nest in the soil [47], and their offspring grow and mature through winter, emerging as new adults the following year. When preparing wildflower beds for planting, tillage is a standard preparation practice [20,48], and research is needed to determine if soil disturbance affects floral resource provision.
The objectives of this research were to (1) determine how the bed preparation method impacts the establishment, growth, and flowering of 15 Southeastern US native forb species planted from plugs, (2) evaluate the effectiveness of the bed preparation method for weed control, and (3) assess the bed preparation method’s impact on arthropod abundance and diversity.
2. Materials and Methods
2.1. Time of Study and Field Location
This study extended over three consecutive years during the growing seasons of 2020, 2021, and 2022. The timing of specific practices is described below. The study was conducted in a peri-urban environment twenty-four miles south of Atlanta, Georgia, on property owned by Trilith Studios (461 Sandy Creek Rd, Fayetteville, GA 30214, Fayette County, USA; 33.4731086, −84.5099865; USDA Plant Hardiness zone 8a). The site was previously used as rangeland and has remained fallow for at least two decades. The study site was an open field that extended one hundred feet at a slope of less than 5° towards a major roadway. Soil type at the experiment site is classified as a piedmont series: fine, mixed, superactive, thermic Udertic Argiustolls. Testing determined that the site’s soil was composed of clay (sand 40%, silt 20%, clay 40%) with a pH of 4.99 and organic matter (2.24%), carbon (0.28%), and nitrogen (0.02%).
2.2. Experimental Design, Plot Layout, and Treatments
The experiment was designed as a strip-plot experiment with two factors, bed preparation method and plant species, with three replicate blocks and eight experimental treatment plots per block. In each block, four plots were prepared via tillage and dazomet (commercial product: Basamid G®, active ingredient: dazomet 99%, AMVAC Chemical Corporation, Newport Beach, CA, USA), while the other four plots were left untilled and treated with glyphosate (commercial product: Ranger Pro®, active ingredient: glyphosate 41%, Southern Agricultural Insecticides, Inc., Palmetto, FL, USA). Each treatment is described in detail further below. Experimental plots contained two randomly planted subsamples of 15 different plant species for a total of 30 plants per plot. Experimental plots measured 13 × 13 feet (3.96 m × 3.96 m) and were separated on all sides by a five-feet (1.52 m) buffer. The study site was approximately 0.16 acres.
2.3. Bed Preparation
To prepare seedbeds for planting, no-till plots were treated with glyphosate on 3 separate occasions. Initially, a treatment solution containing 1.5% glyphosate was applied until runoff to the experimental area with a hand-pressurized backpack sprayer (Lesco 61840, Cleveland, OH, USA). After observing that the first glyphosate application was relatively ineffective (especially on Rubus sp.), the glyphosate concentration was increased from 1.5% to 5% and two additional applications were carried out with the same equipment. In the tilled plots, a tractor (John Deere 4600, Moline, IL, USA) with a tiller attachment (Land Pride RTA3576, Salina, KS, USA) traveled in the northeast–southwest direction and roto-tilled plots to a depth of six inches. Dazomet was then applied at a rate of 400 lbs·A−1. The dazomet-treated plots were then tilled in the same direction at the same depth to incorporate the dazomet into the soil and then packed with a roller (Brouwer BTR-30, Kesmac Inc., Keswick, ON, Canada). A one-inch rainfall event was stimulated using water hoses to properly activate the fumigant. To ensure that each plot received one inch of water, two rain gauges (AcuRite, Lake Geneva, WI, USA) were randomly placed throughout each plot and irrigation ceased once each rain gauge was filled to the one-inch mark. Once the dazomet had been incorporated, compacted, and activated, a compact utility loader (Toro Dingo TX-1000 Wide Track Vertical Lift, Bloomington, IN, USA) with a trencher attachment (Toro, Bloomington, IN, USA) dug a six-inch deep trench around the treated plots. Plastic sheeting (HDX 6 mil clear plastic sheeting) was then laid over the plots, and the edges of the sheeting were buried by hand in the trench. This plastic sheeting was sealed in the dazomet and was removed after twelve days, after which trenches were backfilled.
2.4. Plant Material
Plant material was grown from seeds purchased from Prairie Moon Nursery (Winona, MN, USA). Plants were grown in a greenhouse at the University of Georgia-Griffin campus, USA. Standard protocols for dormant seed and subsequent seedling establishment were followed. Herbaceous perennial species were selected for the following reasons: native status (Georgia and/or SE region), purported pollinator value, suitability for growth in open areas, tolerance of wet to dry soil conditions, and bloom time (Table 1).
Planting occurred on 28 September 2020. Individual plants were transplanted in a triangular pattern on thirty-inch centers in 10–12-inch-deep holes dug by a gas-powered auger equipped with a six-inch drill bit (Stihl, Waiblingen, Germany). The spacing and pattern of planting took into consideration plant size at maturity and industry standards for herbaceous plants. A total of 720 plants were manually planted. Following planting, irrigation was applied by water hoses attached to a water truck on three separate dates to ensure survival of the transplants. A one-inch irrigation event was simulated in each plot and measured with rain gauges. No supplemental irrigation was applied following the initial establishment phase. Natural precipitation over the study period totaled 48.6 inches (October 2020 through August 2021). Additionally, the average daily high was 71.6 °F and the average daily low was 51.0 °F, with the lowest record temperature (21.6 °F) occurring on 17 February 2021 and the highest record temperature (94.0 °F) occurring on 29 July 2021.
The soil was neither amended during or after planting with nutrients or organic matter (as opposed to a typical practice in garden settings), following recommendations for native perennial plantings in prairie and other naturalized settings [49]. All spontaneously occurring vegetation was allowed to grow in the plots, and the buffer zone was not mowed. Surrounding vegetation consisted of naturally recruited forbs that are typical of disturbed sites and were allowed to grow freely.
2.5. Insect-Trapping Methods
To assess overall arthropod community, two types of traps were deployed. Flying insects were trapped using a pan trap consisting of three 12-ounce colored plastic bowls (white, yellow, and blue) secured to a metal frame and were suspended 12 inches off the ground. Deploying three colored bowls [50] is a standard trapping method as different flying insects (i.e., bees, wasps) are more attracted to one color compared to others, especially bowls colored yellow [51]. Pitfall traps were used for insects and arthropods that are active on the soil surface. Each pitfall trap consisted of two nested plastic cups (16 oz). The outer cup had no holes while the inner one was fitted with a 1-inch × 1-inch mesh screen to allow excess water to drain to prevent trapped specimens from washing out. The trap was placed in a hole dug in the middle of the plot. A foam plate was then secured over the trap to prevent rainwater from directly entering the trap. The trap was then filled with soapy solution (10 mL of Tween®20 in 3.79 L water) to break surface tension and facilitate trapping. Both trap types were deployed for three weeks from mid-July till the first week of August 2021. Each plot contained either a pitfall or a pan trap. Traps were randomly assigned to each replicate plot for a total of 24 traps. Traps were emptied once a week, and their contents were transferred to a lab where they were rinsed in running water and placed in labeled containers in 90% ethyl alcohol for preservation.
Insect identification was carried out using standard taxonomic keys (iNaturalist.org) under a stereoscope with a digital camera attachment. The following broad groups were recognized and counted: bees, wasps, Parasitica (broad group encompassing parasitic and parasitoid wasps [52]), flies, butterflies and moths, beetles, herbivores (i.e., thrips, aphids, barklice, truebugs), springtails, crickets, and spiders. Although ants were evident, they were not included in the counts as they are a superorganism, and their presence indicates that a nest(s) is in proximity.
2.6. Data Measurements
2.6.1. Plants
From March to August 2021, bi-weekly measurements of each species were conducted to include plant emergence, plant height (H), plant width (W, two measurements, widest and perpendicular to it), and number of blooms. Growth index (GI) was calculated according to the formula H + ((W1 + W2)/2)/2 [53]. The following were counted as a single bloom: umbel inflorescence (i.e., Allium and Zizia), compound umbel (topmost umbel, i.e., Asclepias), terminal spike (i.e., Agastache and Salvia), and terminal branched inflorescence with multiple blooms per branch (2–5 mm) (i.e., Penthorum). For all Asteraceae species, each capitulum was counted.
2.6.2. Spontaneous Weeds
Weed abundance and coverage were calculated using a 1 m2 PVC quadrat that was randomly tossed within each plot once a week for the duration of the study. If the quadrat landed on a planted wildflower, the quadrat was moved aside and excluded from measurement. Within each quadrat, weeds were counted, and a photograph was taken at noon on a clear day. This was necessary to ensure high contrast and strong color in the digital photograph (minimizing shadows, which could interfere with accurate digital measurements). To assess weed coverage, digital images of the quadrat were processed in Adobe Photoshop Creative Suite® (Adobe, San Jose, CA, USA) using pixel color discrimination. Pixels with a green color and hue were selected by the software, which then produced a count. The value was then recorded in Microsoft Excel® (Microsoft Corporation, Redmond, WA, USA) and statistically analyzed. Although visual observations and number of plants were noted throughout the study period, only data recorded at the end of the study in August 2021 were analyzed and presented.
2.7. Statistical Analysis
Data were analyzed using SAS® v. 9.4 software (SAS Institute, Cary, NC, USA) as follows. Growth index was analyzed using PROC MIXED and log link transformation for normally distributed data. Number of blooms was analyzed using PROC GLIMMIX and log link transformation for count data with Poisson distribution. The model included bed preparation and species as main effects and their one-way interaction, and block and replication, as a random effect. Post hoc mean separation was performed using the Tukey–Kramer test, and significance was determined at the p ≤ 0.05 level. To determine the effect of bed preparation, data on the number of weeds and weed coverage were analyzed with a t-test. Arthropod data were analyzed using a generalized linear mixed model (PROC GLIMMIX) with negative binomial distribution and log link. Back-transformed data are reported. Post hoc mean separation was performed using lsmeans and Tukey–Kramer test, and significance was determined at p ≤ 0.05.
3. Results
3.1. Plant Performance
Bed preparation, plant species, and their interaction were highly significant (Table 2), with growth index and bloom count significantly higher under tillage + dazomet (GI: 47.4, bloom number: 9.4) compared to non-tillage with glyphosate bed preparation (GI: 36.5, bloom number: 5.2).
Under tillage + dazomet, the top three performing species in terms of growth index were S. pilosum, V. angustifolia, and B. asteroides. Similarly, the first two species had the most blooms, but R. fulgida exhibited a higher bloom count than V. angustifolia (Table 3). Although P. sedoides had the lowest GI, it produced more blooms than several other species. The lowest bloom count was observed in A. cernuum. Under non-tillage + glyphosate, the top three performing species in terms of growth index were also S. pilosum, V. angustifolia, and B. asteroides. The first two species had the most blooms, but R. fulgida exhibited a higher bloom count than V. angustifolia. Under this bed preparation treatment, P. sedoides had the lowest growth index (but not significantly different than A. cernuum); however, its bloom count was higher than that of other species with higher growth indices. The lowest bloom number was seen in A. cernuum.
When averaged over both bed treatments, the same trends were observed with respect to the top four performing species, S. pilosum, V. angustifolia, B. asteroides, and R. fulgida (Figure 1).
3.2. Weed Control
Bed preparation had a significant impact on weed species, number, and coverage within the plots, with both weed number and weed coverage being greater in plots that were left untilled and prepared with glyphosate (Table 4 and Figure 4). No-till plots contained more than four times as many individual weeds than their tilled counterparts. The weed species present included blackberry (Rubus spp.), crabgrass (Digitaria spp.), bracted plantain (Plantago aristata), goldenrod (Solidago spp.), and little bluestem (Schizachyrium scoparium), among others.
3.3. Arthropod Presence
Bed preparation had no effect on insect abundance (F = 0.2, p = 0.65), while insect group was highly significant (F = 40.3, p < 0.0001). The interaction effect of bed preparation and insect group was not significant (F = 0.92, p = 0.25). A total of 2529 arthropods were collected in pan and pitfall traps over a period of 3 weeks (Table 5). The most abundant group was flies, closely followed by thrips; both groups had significantly higher counts compared to other groups. Following this were bees, spiders, Parasitica, and truebugs. Barklice, aphids, beetles, wasps, and springtails had significantly fewer numbers than the previous groups. Butterflies/moths and crickets exhibited the lowest numbers.
4. Discussion
We found sufficient evidence to support our hypotheses that plant growth would be impacted by the bed preparation method used and that weed pressure would be reduced with the application of dazomet. Plants grown in beds prepared with tillage and dazomet were larger, produced more blooms, and faced less weed competition compared to their counterparts grown in beds left untilled and treated with glyphosate. The top performing species were S. pilosum, V. angustifolia, B. asteroides, and R. fulgida.
We found that some species were more impacted by bed preparation than others, with the growth index in non-tilled beds being reduced by as much as 55% compared to their tilled counterpart (A. foeniculum). The remaining species exhibited the following percent reduction (in descending order): H. autumnale, 40, B. asteroides, 31, V. angustifolia, 31, A. cernuum, 30, A. virginiana, 21, E. yuccifolium, 20, S. pilosum, 16, S. lyrata, 16, R. fulgida, 13, S. albidum, 12, P. sedoides, 10, A. tuberosa, 9, S. laeve, 8, and Z. aptera, 3.
Since both bed preparation treatments received the same amount of natural irrigation, soil compaction could be the most likely factor causing growth reduction. However, even in the non-tilled beds, soil was loosened to a depth of 10 inches during planting.
All 15 species bloomed, and as expected, their combined phenology spanned March through September. This factor, along with the finding that the plantings attracted an abundant and diverse arthropod community, leads us to conclude that the 15 species can be recommended for provision of season-long floral resources. Although a second year of growth information would be desirable, we were only able to collect data in the spring of 2022 because the entire research site was converted to a parking lot. However, data from two months, April and early May, indicated that plants were showing good growth and, in the case of asters, even spreading three times beyond the ground coverage of the previous year).
Dazomet was effective in suppressing weeds more than eighteen months after its initial application (March 2020). Weed counts were four times greater and weed coverage was more than doubled in no-till and glyphosate plots. Dazomet, which produces methyl-isothiocyanate (MITC), formaldehyde, hydrogen sulfide, and carbon disulfide when activated, degrades in water in less than two days and in a few hours in biologically active soils [54]. Dazomet applications initially sterilize weed seeds, pathogens, insect, and nematodes found in soil [54], and in many cropping systems, weeds and pathogens are inadvertently re-introduced to dazomet-treated sites through either runoff, wind, or human, animal, and insect activity [55]. Researchers often recommend that growers use chemical pesticides and integrated pest management (IPM) programs after applying dazomet to ensure that pest populations are monitored and suppressed [55]. Our study lacked any post-planting chemical, mechanical, or biological weed control methods. Regardless, beds treated with dazomet had fewer weeds, less weed cover, larger wildflower growth, and their plants produced more blooms.
There are several limitations to consider. First, dazomet is a restricted use pesticide and can only be purchased and applied by those possessing the necessary credentials; thus, dazomet is not appropriate for home and garden settings. Second, tillage may damage soil structure [56], leading to erosion and soil runoff. Dazomet can only be used if incorporated into the soil by tillage. Third, our findings contradict previous research; a study on cropland in Virginia found that wildflowers were more successful in beds that were left untilled and treated with glyphosate compared to beds that were rotary power-tilled [48]. The differences in our results might be explained by the use of dazomet in our tilled beds, but further research is needed to examine the residual effects of both dazomet and tillage. Finally, in roadside plantings and other areas, site accessibility might be challenging because applying dazomet demands greater labor and equipment inputs than glyphosate; if a tractor is unable to reach a site, dazomet cannot be used.
This study explored whether bed preparation (tillage and dazomet versus non-tillage and glyphosate) of perennial wildflower plantings impacts arthropod abundance and diversity. Even though plant growth and flowering were significantly lower in beds prepared with no-till and glyphosate treatments, the bed preparation method did not impact the number of insects that visited the plots where the wildflowers were planted. Although this result is unexpected in light of the fact that no-till and glyphosate plots contained fewer blooms, it is encouraging that the insect abundance was comparable to that of tilled plots where plant growth was more robust, indicating that despite diminished plant growth, insects are present. One possible explanation for our findings is that the distance between the individual plots (18 feet between traps) may not have allowed for differences to be detected between plot treatments as flying insects typically have a larger range of flight. For the top three pollinator groups (i.e., bees, wasps, butterflies and moths), the difference between tilled (bees, wasps, butterflies and moths = 122) and non-tilled (bees, wasps, butterflies and moths = 113) plots was small.
With respect to the insect diversity, arthropods from ten taxonomic orders were detected (flies, Order: Diptera; thrips, Order: Thysanoptera; wasps and bees, Order: Hymenoptera; spider, Order: Aranea; truebugs, Order: Hemiptera; barklice, Order: Psocoptera; beetles, Order: Coleoptera; springtails, Order: Collembola; butterflies and moths, Order: Lepidoptera; crickets, Order: Orthoptera). To the best of our knowledge, this is one of few reports exploring not only flying insects but also ground-dwelling arthropods (i.e., springtails, ground beetles, crickets, spiders).
Flies and thrips were the most predominant group, with the number recorded being four and a half times higher than the number of bees. Flies, and in particular long-legged flies (which were the most observed species in the present study), are typically present in high numbers on flowers [30], and often found resting on foliage. They are considered pollinators as well as biological control agents as adults feed on soft-bodied arthropods such as aphids, springtails, and many insect eggs [57]. While thrips have typically been assigned to the category of “pests”, recent studies have claimed that thrips are beneficial as “minute” pollinators and can significantly contribute to the reproductive success of native plants [58].
Bees and wasps were evident in the traps (bees = 177, wasps = 44), and various lower taxa, such as sweat bees, bumble bees, leaf-cutting bees, long-horned bees, scoliid wasps, and potter wasps, were also present. Of particular importance was the presence of insects in the group Parasitica. Parasitica is a broad group that encompasses parasitic and parasitoid wasps, which use other insects (many plant pests) as hosts, depositing eggs which develop inside the host insect and kill the host when mature parasitoid wasps hatch [52]. Parasitoid wasps are important biological control agents, and their presence in high numbers is encouraging, further demonstrating that the plants were able to sustain a diverse arthropod community.
A study conducted in 2013 and 2014 at the University of Griffin, GA Research and Education Garden, used a comparable number of pan traps placed in replicate plots planted with flowering forbs [30]. The researchers collected a total of 214 bees between 25 June 2013 and 4 August 2013, and 25 June 2014 and 4 August 2014. In our study, we collected a comparable number of bees in just a single year with a shorter collection period.
Although there were significantly fewer butterflies and moths compared to bees and wasps (the other two pollinator groups), this variation could be explained by our trapping methods. Passive trapping methods, such as pan and pitfall traps, are highly effective for small- and medium-sized hymenopterans but less effective for larger lepidopterans [52].
Truebugs (Order: Hemiptera) comprise numerous phytophagous species but have many natural enemies, such as assassin bugs and ambush bugs. Certain families of leaf bugs, leafhoppers, treehoppers, and seed bugs are typically found in high numbers on flowering forbs [30,59]. A total of 105 hemipterans were collected in the current study, a relatively low amount. Once more, this variation could be explained by our trapping methods. Sweep netting typically results in the capture of more hemipterans [35]. Similarly, beetles (Order: Coleoptera) were recorded in relatively low numbers. They too could have been captured in higher numbers if a sweep net were used.
Spiders were captured in both pan traps and pitfall traps; the latter method is typically used to collect this taxon. As spiders are predators and consume other insects, their presence is evidence of activity among higher trophic levels. The fact that 158 spiders were trapped also points to the presence of a reasonably abundant arthropod community.
Our study was able to provide empirical data on how 15 plug-grown native wildflowers performed under punishing climactic conditions—poor soil and little water. It is encouraging that all 15 species performed as well as they did under non-tilled conditions. At the time of the study’s conclusion, 91% of the plants in the till + dazomet plots appeared to be alive compared to 85% of plants in the no-till and glyphosate plots [60]. Only E. yuccifolium and A. tuberosa have been previously evaluated when grown as transplants under mesic conditions in Michigan [33]. Based on that study, researchers concluded that the latter should be recommended as a high-value species for pollinators and other beneficial insects.
While planting plugs is more time-consuming than planting seeds, wildflowers planted as plugs might bloom earlier than those planted from seed, and as shown by our research, are able to survive warmer summers without supplemental irrigation. Their performance in future years needs to be further researched, but at this time, they appear to be a viable tool for rapid wildflower bed establishment.
Managing customer expectations must be a major educational component of wildflower plantings. Many native wildflower species are not as visually appealing as other more common non-native plant species. Before planting wildflower beds, land managers and landscaping companies need to help customers learn to visually accept wildflower spaces which are less orderly and more naturalistic than traditional landscape plantings [61]. Cultural discussions must take place regarding how wildflower plantings look year-long, future maintenance, and their importance to preserving pollinator health. For much of the winter, wildflower plantings lie dormant, lacking presence and color. Major political changes are forcing the scientific community to find new solutions to our current problems, and dazomet will hopefully be a part of those solutions. The industry standard for bed preparation, using glyphosate to clear land of vegetation prior to planting, may not be possible in the near future. Glyphosate is currently approved for use in the European Union (EU) until 15 December 2023, after which a team of global experts will decide whether or not to approve its renewal [62]. Future generations might be unable to purchase or use glyphosate.
5. Conclusions
This study evaluated the effect of bed preparation on wildflower establishment, weed control, and arthropod presence. Growth index and bloom count were significantly higher under tillage + dazomet (GI: 47.4, bloom number: 9.4) compared to non-tillage + glyphosate bed preparation (GI: 36.5, bloom number: 5.2). There were 8.1 weeds per quadrat in the tillage + dazomet treatment compared to 33.8 weeds in the non-tillage + glyphosate treatment. Our results show that dazomet can be used as a tool to prepare wildflower beds for planting. Of the 15 native wildflower species evaluated in this study, each one grew larger, produced more blooms, and had to compete with fewer weeds when grown in beds treated with dazomet. The top performing species were S. pilosum, V. angustifolia, B. asteroides, and R. fulgida (GI, bloom count: 77.8, 22.7; 58.1, 5.1; 51.6, 10.2; 46.9, 3.0, respectively). Sisyrinchium albidum, R. fulgida, S. laeve, and S. pilosum were in bloom for two months, while the rest of the species bloomed for a month. Additionally, all species bloomed less than one year after being transplanted as a plug, demonstrating that this method of starting wildflower habitat is a viable option for land managers, landscaping companies, and civic entities looking to quickly establish plantings. Despite not receiving supplemental fertilizer, organic matter, or irrigation, over 90% of the plants in the no-till with dazomet plots survived and were able to attract and sustain pollinator and beneficial insect populations.
Author Contributions
Conceptualization, S.V.P.; methodology, S.V.P. and M.C.; validation, S.V.P. and M.C.; formal analysis, J.S.S.; investigation, J.S.S.; resources, S.V.P. and J.S.S.; data curation, S.V.P.; writing—original draft preparation, J.S.S.; writing—review and editing, J.S.S., S.V.P. and M.C.; visualization, J.S.S. and S.V.P.; supervision, S.V.P.; project administration, S.V.P.; funding acquisition, S.V.P. and J.S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- Brereton, T.M.; Warren, M.S.; Roy, D.B.; Stewart, K. The changing status of the Chalkhill Blue butterfly Polyommatus coridon in the UK: The impacts of conservation policies and environmental factors. J. Insect Conserv. 2008, 12, 629–638. [Google Scholar] [CrossRef]
- Modiba, R.V.; Joseph, G.S.; Seymour, C.L.; Fouché, P.; Foord, S.H. Restoration of riparian systems through clearing of invasive plant species improves functional diversity of Odonate assemblages. Biol. Conserv. 2017, 214, 46–54. [Google Scholar] [CrossRef]
- Carrié, R.; Ekroos, J.; Smith, H.G. Organic farming supports spatiotemporal stability in species richness of bumblebees and butterflies. Biol. Conserv. 2018, 227, 48–55. [Google Scholar] [CrossRef]
- Hopwood, J.; Black, S.H.; Lee-Mäder, E.; Charlap, A.; Preston, R.; Mozumder, K.; Fleury, S. Literature Review: Pollinator Habitat Enhancement and Best Management Practices in Highway Rights-of-Way; The Xerces Society for Invertebrate Conservation and ICF International, Federal Highway Administration: Washington, DC, USA, 2015. [Google Scholar]
- Perry, L. Successful Wildflower Meadows; University of Vermont Extension: Burlington, VT, USA, 2005; OH 84. [Google Scholar]
- Easton, V. Wildflower mixes: The good, the bad, the ugly. The Seattle Times, 25 February 2012. [Google Scholar]
- Haaland, C.; Nesbitt, R.E.; Bersier, O.F. Sown wildflower strips for insect conservation: A review. Insect Conserv. Divers. 2011, 4, 60–80. [Google Scholar] [CrossRef]
- Stokstad, E. Pesticides Under Fire for Risks to Pollinators. Science 2013, 340, 674–676. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Christmann, S. Pollinator protection strategies must be feasible for all nations. Nat. Ecol. Evol. 2020, 4, 896–897. [Google Scholar] [CrossRef]
- Bretzel, F.; Pezzarossa, B.; Carrai, C.; Malorgio, F. Wildflower plantings to reduce the management costs of urban gardens and roadsides. Acta Hortic. 2009, 813, 263–270. [Google Scholar] [CrossRef]
- Campbell, J.W.; Kimmel, C.B.; Grodsky, S.M.; Smithers, C.; Daniels, J.C.; Ellis, J.D. Wildflower plantings harbor increased arthropod richness and abundance within agricultural areas in Florida (USA). Ecosphere 2019, 10, e02890. [Google Scholar] [CrossRef] [Green Version]
- Ahern, J.; Barker, A. Roadside wildflower meadows: Summary of benefits and guidelines to successful establishment and management. Transp. Res. Rec. 1992, 1334, 46–53. [Google Scholar]
- Derr, J.F.; Appleton, B.L. Weed control with landscape fabrics. J. Environ. Hortic. 1989, 7, 129–133. [Google Scholar] [CrossRef]
- Corley, W.; Murphy, T.; Reynolds, K. Weed management options for wildflower meadows and beauty spots. SNA Res. Conf. Proc. 1993, 38, 135. [Google Scholar]
- Aldrich, J.H. Factors and Benefits in the Establishment of Modest-Sized Wildflower Plantings: A Review. Nativ. Plants J. 2002, 3, 67–86. [Google Scholar] [CrossRef] [Green Version]
- Skousen, J.; Venable, C. Establishing native plants on newly-constructed and older-reclaimed sites along West Virginia highways. Land Degrad. Dev. 2008, 19, 388–396. [Google Scholar] [CrossRef]
- Angelella, G.M.; Stange, L.; Scoggins, H.L.; O’rourke, M.E. Pollinator refuge establishment and conservation value: Impacts of seedbed preparations, seed mixtures, and herbicides. Hortscience 2019, 54, 445–451. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.R.; McCullough, P.E.; Shilling, D.G. Native plant establishment on georgia roadsides. Agron. J. 2015, 107, 990–996. [Google Scholar] [CrossRef]
- Kanissery, R.; Gairhe, B.; Kadyampakeni, D.; Batuman, O.; Alferez, F. Glyphosate: Its Environmental Persistence and Impact on Crop Health and Nutrition. Plants 2019, 8, 499. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.P.; Peden, J.M.M.; Dick, R.E. Glyphosate tolerance and utilization by the microflora of soils treated with the herbicide. Appl. Microbiol. Biotechnol. 1988, 29, 511–516. [Google Scholar] [CrossRef]
- Gilreath, J.P.; Santos, B.M. Efficacy of methyl bromide alternatives on purple nutsedge (Cyperus rotundus) control in tomato and pepper. Weed Technol. 2004, 18, 341–345. [Google Scholar] [CrossRef]
- Fennimore, S.A.; Haar, M.J.; Goodhue, R.E.; Winterbottom, C.Q. Weed Control in Strawberry Runner Plant Nurseries with Methyl Bromide Alternative Fumigants. Hortscience 2008, 45, 1495–1500. [Google Scholar] [CrossRef] [Green Version]
- Czarnota, M.A. Georgia Pest Management Handbook; Commercial Edition; University of Georgia: Athens, GA, USA, 2016. [Google Scholar]
- Environmental Protection Agency. Climate Change Indicators: U.S. and Global Precipitation; United States Environmental Protection Agency: Washington, DC, USA, 2021. [Google Scholar]
- Norcini, J.G.; Aldrich, J.H. Establishment of Native Wildflower Plantings by Seed; ENH1046; University of Florida: Gainesville, FL, USA, 2004. [Google Scholar]
- Milstein, G.P. The uses and potential of wildflower seed in landscaping. In Flower Seeds: Biology and Technology; CABI Publishing: Columbus, OH, USA, 2005; pp. 39–51. [Google Scholar]
- Norcini, J.G.; Aldrich, J.H.; Halsey, L.A.; Lilly, J.G. Seed source affects performance of six wildflower species. Proc. Fla. State Hort. Soc. 1998, 111, 4–9. [Google Scholar] [CrossRef]
- Harris, B.A.; Braman, S.K.; Pennisi, S.V. Influence of plant taxa on pollinator, butterfly, and beneficial insect visitation. Hortscience 2016, 51, 1016–1019. [Google Scholar] [CrossRef] [Green Version]
- Dantas, B.F.; Moura, M.S.B.; Pelacani, C.R.; Angelotti, F.; Taura, T.A.; Oliveira, G.M.; Bispo, J.S.; Matias, J.R.; Silva, F.F.S.; Pritchard, H.W.; et al. Rainfall, not soil temperature, will limit the seed germination of dry forest species with climate change. Oecologia 2020, 192, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Whittinghill, L.J.; Rowe, D.B.; Ngouajio, M.; Cregg, B.M. Evaluation of nutrient management and mulching strategies for vegetable production on an extensive green roof. Agroecol. Sustain. Food Syst. 2016, 40, 297–318. [Google Scholar] [CrossRef]
- Gibson, D.R.; Rowe, L.; Isaacs, R.; A Landis, D. Screening Drought-Tolerant Native Plants for Attractiveness to Arthropod Natural Enemies in the U.S. Great Lakes Region. Environ. Èntomol. 2019, 48, 1469–1480. [Google Scholar] [CrossRef]
- Campbell, J.W.; Smithers, C.; Irvin, A.; Kimmel, C.B.; Stanley-Stahr, C.; Daniels, J.C.; Ellis, J.D. Trap nesting wasps and bees in agriculture: A comparison of sown wildflower and fallow plots in florida. Insects 2017, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Harris, B.A.; Poole, E.; Pennisi, S.V. Impact of biostimulant and cultural factors on whorled mountain mint Pycnanthemum verticillatum (Michx.) Pers. var. pilosum (Nutt.) Cooperr.: Growth performance and assessment of arthropod visitation. Nat. Plants J. 2022, 23, 97–114. [Google Scholar] [CrossRef]
- Braman, S.K.; Pennisi, S.V.; Fair, C.G.; Quick, J.C. Pollinator cultivar choice: An assessment of season-long pollinator visitation among coreopsis, aster, and salvia cultivars. Front. Sustain. Cities 2022, 4, 988966. [Google Scholar] [CrossRef]
- Kalaman, H.; Wilson, S.B.; Mallinger, R.E.; Knox, G.W.; Kim, T.; Begcy, K.; van Santen, E. Evaluation of native and nonnative ornamentals as pollinator plants in Florida: II. Floral resource value. HortScience 2021, 57, 137–143. [Google Scholar] [CrossRef]
- Palmersheim, M.C.; Schürch, R.; O’rourke, M.E.; Slezak, J.; Couvillon, M.J. If you grow it, they will come: Ornamental plants impact the abundance and diversity of pollinators and other flower-visiting insects in gardens. Horticulturae 2022, 8, 1068. [Google Scholar] [CrossRef]
- Blaauw, B.R.; Isaacs, R. Larger wildflower plantings increase natural enemy density, diversity, and biological control of sentinel prey, without increasing herbivore density. Ecol. Èntomol. 2012, 37, 386–394. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, G. Formation, characteristics and eco-environmental implications of urban soils—A review. Soil Sci. Plant Nutr. 2015, 61 (Suppl. S1), 30–46. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Zhang, G. Differences in shallow soil temperatures at urban and rural areas. J. Eng. Geol. 2012, 20, 58–65. [Google Scholar]
- Schellenberg, M.P.; Biligetu, B.; Lamb, E.G.; Mischkolz, J.M. World interest in diverse native plant stands. In PXXII International Grassland Congress Proceedings; University of Kentucky: Lexington, KY, USA, 2022. [Google Scholar]
- Bang, C.; Faeth, S.H. Variation in arthropod communities in response to urbanization: Seven years of arthropod monitoring in a desert city. Landsc. Urban Plan. 2011, 103, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Sutter, L.; Albrecht, M.; Jeanneret, P. Landscape greening and local creation of wildflower strips and hedgerows promote multiple ecosystem services. J. Appl. Ecol. 2017, 55, 612–620. [Google Scholar] [CrossRef]
- Mody, K.; Lerch, D.; Müller, A.-K.; Simons, N.K.; Blüthgen, N.; Harnisch, M. Flower power in the city: Replacing roadside shrubs by wildflower meadows increases insect numbers and reduces maintenance costs. PLoS ONE 2020, 15, e0234327. [Google Scholar] [CrossRef]
- Ullmann, K.S.; Meisner, M.H.; Williams, N.M. Impact of tillage on the crop pollinating, ground-nesting bee, Peponapis pruinosa in California. Agric. Ecosyst. Environ. 2016, 232, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.; Carril, O. The Bees in Your Backyard; Princeton University Press: Princeton, NJ, USA, 2016. [Google Scholar]
- Angelella, G.M.; O’rourke, M.E. Pollinator habitat establishment after organic and no-till seedbed preparation methods. Hortscience 2017, 52, 1349–1355. [Google Scholar] [CrossRef] [Green Version]
- Jordan, S.F.C.; Jessa, K.; Gill, K.; Hopwood, J.; Fowler, J.; Lee-Mader, E.; Vaughan, M. Wildflower Establishment, Organic Site Preparation Methods; Xerces Society for Invertebrate Conservation: Portland, OR, USA, 2016. [Google Scholar]
- Parys, K.A.; Esquivel, I.L.; Wright, K.W.; Griswold, T.; Brewer, M.J. Native Pollinators (Hymenoptera: Anthophila) in Cotton Grown in the Gulf South, United States. Agronomy 2020, 10, 698. [Google Scholar] [CrossRef]
- Harris, B.; Braman, S.; Pennisi, S. Pan trap designs for monitoring pollinators and other beneficial insects in conservation gardens. J. Èntomol. Sci. 2017, 52, 9–14. [Google Scholar] [CrossRef]
- Triplehorn, C.A.; Johnson, N.F. Chapter 28: Order Hymenoptera. In Borror and DeLong’s Introduction to the Study of Insects, 7th ed.; Brooks/Cole, Cengage Learning: Boston, MA, USA, 2019. [Google Scholar]
- Irmak, S.; Haman, D.Z.; Irmak, A.; Jones, J.W.; Campbell, K.L.; Crisman, T.L. Measurement and Analyses of Growth and Stress Parameters of Viburnum odoratissimum (Ker-gawl) Grown in a Multi-pot Box System. Hortscience 2004, 39, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Shaner, D.C. Herbicide Handbook, 10th ed.; Weed Science Society of America: Westminster, CO, USA, 2014. [Google Scholar]
- Lyons, M.T. Composts, Biofumigation, Dazomet, and Integrated Treatments Influence Southern Blight, Yield and Microbial Community Composition in Plasticulture Tomato Production; The University of Tennessee: Knoxville, TN, USA, 2003. [Google Scholar]
- Nunes, M.R.; Karlen, D.L.; Moorman, T.B. Tillage intensity effects on soil structure indicators—A US meta-analysis. Sustainability 2020, 12, 2071. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, M.M. Chapter 8: Predatory Flies. In Good Garden Bugs: Everything You Need to Know About Beneficial Predatory Insects; Quarry Books: Beverly, MA, USA, 2015. [Google Scholar]
- Eliyahu, D.; McCall, A.C.; Lauck, M.; Trakhtenbrot, A.; Bronstein, J.L. Minute pollinators: The role of thrips (Thysanoptera) as pollinators of pointleaf manzanita, Arctostaphylos pungens (Ericaceae). J. Pollinat. Ecol. 2015, 16, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Poythress, J.C.; Affolter, J.M. Ecological Value of Native Plant Cultivars versus Wild-Type Native Plants for Promoting Hemipteran Diversity in Suburban Areas. Environ. Èntomol. 2018, 47, 890–901. [Google Scholar] [CrossRef]
- Shelton, J. Does Bed Preparation Impact Native Wildflower Establishment? Cost Analysis and Implications for Biodiversity; University of Georgia: Athens, GA, USA, 2021. [Google Scholar]
- Huber, G. Understanding Perceptions to Improve the Success and Acceptance of Insect Pollinator Habitat in Public Spaces; University of Georgia: Athens, GA, USA, 2020. [Google Scholar]
- European Union Commission. Available online: https://food.ec.europa.eu/plants/pesticides/approval-active-substances/renewal-approval/glyphosate_en#:~:text=The%20Appeal%20Committee%20also%20did,glyphosate%20until%2015%20December%202023 (accessed on 1 May 2021).
Figure 1.
Performance of 15 species of native perennials growth at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. Data pooled from both bed treatments. Species arranged from highest to lowest values for growth index. Columns with the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level.
Figure 1.
Performance of 15 species of native perennials growth at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. Data pooled from both bed treatments. Species arranged from highest to lowest values for growth index. Columns with the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level.

Figure 2.
Photos of the 15 species of native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. (A). Agastache foeniculum, (B). Eryngium yuccifolium, (C). Boltonia asteroides, (D). Allium cernuum, (E). Sisyrinchium albidum, (F). Anemone virginiana, (G). Helenium autumnale, (H). Symphyotrichum laeve, (I). Asclepias tuberosa, (J). Penthorum sedoides, (K). Salvia lyrata, (L). Rudbeckia fulgida, (M). Vernonia angustifolia, (N). Zizia aptera, (O). Symphyotrichum pilosum.
Figure 2.
Photos of the 15 species of native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. (A). Agastache foeniculum, (B). Eryngium yuccifolium, (C). Boltonia asteroides, (D). Allium cernuum, (E). Sisyrinchium albidum, (F). Anemone virginiana, (G). Helenium autumnale, (H). Symphyotrichum laeve, (I). Asclepias tuberosa, (J). Penthorum sedoides, (K). Salvia lyrata, (L). Rudbeckia fulgida, (M). Vernonia angustifolia, (N). Zizia aptera, (O). Symphyotrichum pilosum.

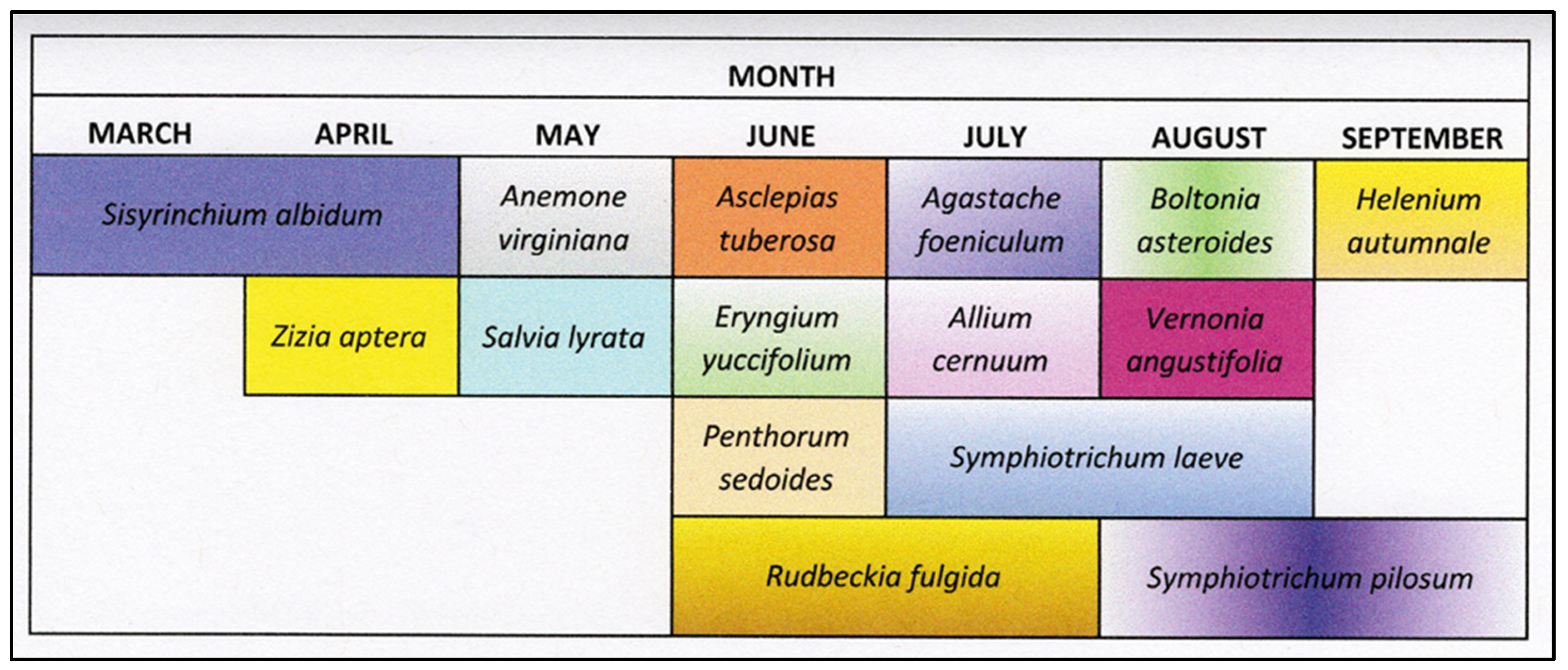
Figure 3.
Bloom period of the 15 native perennials evaluated at the Trilith research site (Fayetteville, GA, USA). Plants evaluated from March to September 2021. Individual box color corresponds to flower color.
Figure 3.
Bloom period of the 15 native perennials evaluated at the Trilith research site (Fayetteville, GA, USA). Plants evaluated from March to September 2021. Individual box color corresponds to flower color.

Figure 4.
Quadrat views of weed cover within individual replicate plots planted with 15 native perennial species. All three blocks are represented. Left column (A,C,E)—bed preparation: tillage with dazomet; right column (B,D,F)—bed preparation: no-till with glyphosate.
Figure 4.
Quadrat views of weed cover within individual replicate plots planted with 15 native perennial species. All three blocks are represented. Left column (A,C,E)—bed preparation: tillage with dazomet; right column (B,D,F)—bed preparation: no-till with glyphosate.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Species planted at the Trilith Studio (Fayetteville, GA, USA) research site in 2020 and evaluated in 2021. Scientific name and taxonomic information, common name, native range, and blooming period included (according to the Lady Bird Johnson Wildflower Center’s plant database, https://www.wildflower.org/plants/ accessed on 5 July 2023).
Table 1.
Species planted at the Trilith Studio (Fayetteville, GA, USA) research site in 2020 and evaluated in 2021. Scientific name and taxonomic information, common name, native range, and blooming period included (according to the Lady Bird Johnson Wildflower Center’s plant database, https://www.wildflower.org/plants/ accessed on 5 July 2023).
| Plant Species | Common Name | Native Range | Blooming Period |
|---|---|---|---|
| Agastache foeniculum ((Nutt.) Britton) (Lamiales; Lamiaceae) | anise hyssop | Northeastern United States | June to September |
| Allium cernuum Roth. (Liliales; Liliaceae) | wild-nodding onion | Northeastern United States | June to August |
| Anemone virginiana L. (Ranunculales; Ranunculaceae) | thimbleweed | United States | April to May |
| Asclepias tuberosa L. (Gentianales: Asclepiadaceae) | butterfly weed | Eastern North America | June to August |
| Boltonia asteroides L. (Asterales; Asteraceae) | white boltonia | Eastern North America | August to September |
| Eryngium yuccifolium Michx. (Apiales: Apiaceae) | rattlesnake master | United States | June to September |
| Helenium autumnale L. (Asterales; Asteraceae) | common sneezeweed | Eastern North America | August to October |
| Penthroum sedoides L. (Saxifragales; Penthoraceae) | ditch stonecrop | North America | July to September |
| Rudbeckia fulgida Aiton. (Asterales: Asteraceae) | orange coneflower | Southeastern United States | June to October |
| Salvia lyrata L. (Lamiales; Lamiaceae) | lyre-leaf sage | United States | April to June |
| Sisyrinchium albidum Raf. (Asparagales; Iridaceae) | blue-eyed grass | North America | May to July |
| Symphyotrichum laeve L. (Asterales; Asteraceae) | smooth blue aster | North America | September to October |
| Symphyotrichum pilosum L. (Asterales; Asteraceae) | hairy aster | North America | August to October |
| Vernonia angustifolia Michx. (Asterales; Asteraceae) | narrow-leaf ironweed | Eastern United States | June to September |
| Zizia aptera (A. Gray) Fernald (Apiales; Apiaceae) | meadow zizia | North America | May |
Table 2.
Analysis of variance results for main effects and their interaction on performance of 15 native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021.
Table 2.
Analysis of variance results for main effects and their interaction on performance of 15 native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021.
| Effect | Plant Morphometrics | |||
|---|---|---|---|---|
| Growth Index | Number of Blooms | |||
| F-Value | Pr > F | F-Value | Pr > F | |
| Bed Preparation | 182.4 | <0.0001 | 577.6 | <0.0001 |
| Plant Species | 136.6 | <0.0001 | 591.3 | <0.0001 |
| Bed Preparation × Plant Species | 10.4 | <0.0001 | 20.2 | <0.0001 |
Table 3.
Growth index and number of blooms in the tillage with dazomet treatment of 15 species of native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. Species arranged from highest to lowest values for growth index in the tillage + dazomet treatment. Means followed by the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level. Standard error in parentheses.
Table 3.
Growth index and number of blooms in the tillage with dazomet treatment of 15 species of native perennials grown at Trilith Studio research site (Fayetteville, GA, USA) in 2020 and 2021. Species arranged from highest to lowest values for growth index in the tillage + dazomet treatment. Means followed by the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level. Standard error in parentheses.
| Species | Treatment | |||
|---|---|---|---|---|
| Tillage + Dazomet | Non-Tillage + Glyphosate | |||
| Growth Index | No. of Blooms | Growth Index | No. of Blooms | |
| Symphyotrichum pilosum | 92.7 (3.1) a | 50.0 (3.5) a | 77.8 (3.1) a | 22.7 (1.7) a |
| Vernonia angustifolia | 84.3 (2.3) ab | 16.1 (1.2) d | 58.1 (2.3) b | 5.1 (0.4) d |
| Boltonia asteroides | 75.3 (1.9) b | 28.0 (2.0) b | 51.6 (1.9) bc | 10.2 (0.8) b |
| Eryngium yuccifolium | 53.8 (1.9) c | 7.7 (0.6) g | 42.9 (1.0) cd | 4.2 (0.3) de |
| Agastache foeniculum | 52.8 (2.2) c | 6.0 (0.5) ghi | 24.3 (2.1) gh | 2.9 (0.4) ef |
| Symphyotrichum laeve | 51.0 (2.2) c | 6.8 (0.6) gh | 46.9 (2.4) bcd | 3.0 (0.4) ef |
| Helenium autumnale | 50.3 (2.4) c | 4.0 (0.5) ij | 29.6 (2.4) fgh | 4.0 (0.5) def |
| Rudbeckia fulgida | 46.6 (2.3) cd | 19.3 (1.4) c | 41.3 (2.3) cde | 7.9 (0.6) c |
| Salvia lyrata | 44.1 (2.3) cde | 6.0 (0.5) hi | 37.3 (2.3) def | 3.4 (0.3) ef |
| Anemone virginiana | 32.6 (2.9) defg | 5.5 (0.7) ghij | 25.7 (3.2) efgh | 4.2 (0.7) cdef |
| Aclepias tuberosa | 32.6 (2.3) ef | 9.5 (0.7) f | 29.9 (2.4) efg | 5.1 (0.4) d |
| Zizia aptera | 28.3 (2.4) fg | 5.9 (0.5) hi | 26.6 (2.4) fgh | 4.2 (0.3) de |
| Sisyrinchium albidum | 25.4 (2.3) fg | 12.4 (0.9) e | 21.8 (2.1) gh | 7.5 (0.6) c |
| Allium cernuum | 22.9 (2.3) fg | 3.7 (0.3) ij | 16.5 (2.3) h | 2.5 (0.3) f |
| Penthorum sedoides | 18.9 (2.4) g | 8.2 (0.9) fg | 16.9 (2.4) h | 7.7 (0.7) f |
Table 4.
Effect of bed preparation on weed number and coverage. Means followed by the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level. Standard error in parentheses. Coverage was estimated from digital images and processed with Adobe Photoshop® 2023, using pixel color discrimination. Number indicates green pixels.
Table 4.
Effect of bed preparation on weed number and coverage. Means followed by the same letters are not significant according to the Tukey–Kramer test at the p ≤ 0.05 significance level. Standard error in parentheses. Coverage was estimated from digital images and processed with Adobe Photoshop® 2023, using pixel color discrimination. Number indicates green pixels.
| Bed Preparation | Number of Weeds | Weed Coverage |
|---|---|---|
| Tillage + dazomet | 8.1 (0.9) b | 462.5 (65.7) |
| Non-tillage + glyphosate | 33.8 (3.0) a | 1025.7 (145.7) |
Table 5.
Arthropod abundance in bowl and pitfall traps collected in wildflower plots from mid-July to the first week of August 2021. Values are pooled from both bed treatments and presented according to total number of arthropods, from highest to lowest. Mean separation of insect groups according to Tukey–Kramer test. Means followed by the same letters are not significant at the p ≤ 0.05 significance level. Standard error in parentheses.
Table 5.
Arthropod abundance in bowl and pitfall traps collected in wildflower plots from mid-July to the first week of August 2021. Values are pooled from both bed treatments and presented according to total number of arthropods, from highest to lowest. Mean separation of insect groups according to Tukey–Kramer test. Means followed by the same letters are not significant at the p ≤ 0.05 significance level. Standard error in parentheses.
| Arthropod Order/Group | Number | Mean |
|---|---|---|
| Diptera | 819 | 5.69 (1.22) a |
| Thysanoptera | 812 | 5.64 (0.07) a |
| Hymenoptera: bees | 177 | 1.23 (0.17) bc |
| Aranea | 158 | 1.10 (0.23) b |
| Parasitica | 126 | 0.87 (0.12) bcd |
| Hemiptera | 105 | 0.73 (0.12) bcd |
| Psocoptera | 95 | 0.66 (0.10) ced |
| Hemiptera: aphids | 76 | 0.52 (0.07) fed |
| Coleoptera | 66 | 0.46 (0.07) fed |
| Hymenoptera: wasps | 44 | 0.30 (0.05) feg |
| Collembola | 32 | 0.22 (0.04) fhg |
| Lepidoptera | 14 | 0.10 (0.02) hg |
| Orthoptera | 5 | 0.03 (0.01) h |
| Total number | 2529 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shelton, J.S.; Pennisi, S.V.; Czarnota, M. Effect of Bed Preparation on Native Wildflower Establishment, Weed Control, and Arthropod Presence. Horticulturae 2023, 9, 854. https://doi.org/10.3390/horticulturae9080854
AMA Style
Shelton JS, Pennisi SV, Czarnota M. Effect of Bed Preparation on Native Wildflower Establishment, Weed Control, and Arthropod Presence. Horticulturae. 2023; 9(8):854. https://doi.org/10.3390/horticulturae9080854
Chicago/Turabian StyleShelton, Joseph S., Svoboda V. Pennisi, and Mark Czarnota. 2023. "Effect of Bed Preparation on Native Wildflower Establishment, Weed Control, and Arthropod Presence" Horticulturae 9, no. 8: 854. https://doi.org/10.3390/horticulturae9080854
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.